|
ГЛАВНАЯ
> Вернуться к содержанию
Арктика и Антарктика
Правильная ссылка на статью:
Трегубов О.Д.
Проблемы плейстоценовой фауны: термокарст как фактор исчезновения мамонтов
// Арктика и Антарктика.
2017. № 4.
С. 1-12.
DOI: 10.7256/2453-8922.2017.4.24561 URL: https://nbpublish.com/library_read_article.php?id=24561
Проблемы плейстоценовой фауны: термокарст как фактор исчезновения мамонтов
Трегубов Олег Дмитриевич
кандидат геолого-минералогических наук
ведущий научный сотрудник, Северо-Восточный комплексный научно-исследовательский институт ДВО РАН
689000, Россия, Чукотский автономный округ, г. Анадырь, ул. Энергетиков, 5
Tregubov Oleg Dmitrievich
PhD in Geology and Mineralogy
Leading Scientific Associate, N. A. Shilo North-East Interdisciplinary Scientific Research Institute, Eastern Branch of the Russian Academy of Sciences
689000, Russia, Chukotka Autonomous Okrug, Anadyr, Energetikov Street 5, unit #23

|
tregubov2@yandex.ru
|
|
 |
|
DOI: 10.7256/2453-8922.2017.4.24561
Дата направления статьи в редакцию:
28-10-2017
Дата публикации:
10-01-2018
Аннотация:
Рассматривается аргументация основных гипотез исчезновения крупной плейстоценовой фауны. Выделяются и обсуждаются четыре составляющие проблемы – хронологическая, ландшафтно-климатическая, антропогенная и эволюционная. Анализируется возможность влияния метода датировки на представления о хронологии расцвета и упадка популяций мамонтовой фауны. Обсуждаются различия ландшафтных условий гибели и захоронения ископаемой фауны, временные связи массовой гибели животных с эпохами похолодания и потепления. Оценивается роль антропогенного фактора в исчезновении мамонтов. Особое внимание уделяется лёссам как фактору формирующему среду обитания плейстоценовой фауны. Предлагается несколько оригинальных гипотез вымирания мамонтов, которые основываются на закономерностях эволюции биосферы и теории катастроф. Существенное влияние на деградацию мамонтовой фауны на рубеже плейстоцена и голоцена оказали: а) резкое усиление зональности природной среды с сокращением ареала интразональных ландшафтов; б) нарушение путей сезонной и многолетней миграции плейстоценовых животных, в первую очередь наиболее уязвимых к изменению рациона крупных млекопитающих; в) смешение ресурсно-конкурентных зооценозов, которое привело к деградации пастбищ и нарушению трофических цепей. Ключевая роль в нарушении путей миграции и истощении пастбищ отводится термокарстовой катастрофе.
Ключевые слова:
плейстоценовая фауна, палеозоология, ледниковый цикл, вымирание мамонтов, лёссы, подземные льды, термокарст, палеогеография, тундростепи, едома
Abstract: This article examines the argumentation of the main hypothesis of disappearance of the large Pleistocene fauna. The four components of the problem are being determined and discussed – chronological, landscape-climatic, anthropogenic, and evolutionary. The author analyses the possible effect of the dating method upon the perceptions regarding the chronology of rise and fall of the populations of mammoth fauna, as well as different landscape factors of the destruction and disposal of the fossil fauna, temporal connection of the mass destruction of animals with the eras of climatic cooling and warming. The role of anthropogenic factor of mammoths’ extinction is being evaluated. Special attention is given to the loesses as a factor that forms the habitat of Pleistocene fauna. The article suggest several original hypothesis of mammoths’ extinction that are based on the principles of biospheric evolution and catastrophism. Degradation of mammoth fauna at the turn of Pleistocene and Holocene was significantly affected by the following: rapid increase of zonality of the natural environment with reduction in the area of intrazonal landscapes; disruption of the patterns of seasonal and long-term migration of Pleistocene animals, primarily most vulnerable to changes in ration of the large mammals; amalgamation of the resource-viable zoocenosis that led to degradation of grazing lands and disruption of trophic chain. The key role in disruption of the migration patterns and pasture exhausting is assigned to the thermokarst catastrophe.
Keywords: Pleistocene faune, paleozoology, glacial cycle, extinction mammoths, loess, ground ice, thermokarst, paleogeography, tundra-steppe, yedoma
Введение
За последние два столетия мамонтовая загадка стала своеобразным символом и даже логотипом палеонтологических, палеоботанических, палеографических и археологических исследований позднего плейстоцена. О чем свидетельствует неуменьшающееся число публикаций в научных и научно-популярных изданиях, количество предлагаемых новых гипотез или обоснований уже существующих. Условно в проблеме исчезновения мамонтовой фауны можно выделить хронологическую, ландшафтно-климатическую, антропогенную и эволюционную составляющие. При этом сразу нужно отметить, что два, казалось бы, неразрывно, связанных вопроса об условиях гибели и причинах вымирания мамонтов принципиально различны как по содержанию, так и по существу [1].
Хронология событий мамонтовой эпохи
Возраст находок мамонтовых захоронений позднего плейстоцена по данным радиоуглеродных датировок колеблется в интервале 60-10 тыс. лет, а самые поздние датировки ископаемых остатков 7,6-3,7 тыс. лет получены по останкам острова Врангеля (рис. 1). Большая часть обнаружений «молодых» останков мамонтовой фауны приурочено к континентальным отложениям конца плейстоцена - 10-14 тыс. лет [2]. При этом надежный предельный возраст радиоуглеродной датировки порядка органического вещества - 40-60 тыс. лет. Считается, что мамонты появились в конце плиоцена – начале плейстоцена около 1,5-2 млн. лет назад и уже 350-250 тыс. лет назад колонизировали Евразию и Северную Америку, достигнув расцвета в начале предпоследнего оледенения - 60-70 тыс. лет назад [3].
Как видно общим в хронологии событий является рубеж 60 тыс. лет, соответствующий: а) первому максимуму зырянского (валдайского, вюрмского) оледенения; б) нижнему пределу радиоуглеродного метода датировки; в) максимуму популяции шерстистого мамонта; г) самому раннему времени массовых находок захоронений мамонтовой фауны. Иными словами наибольшее число и плотность захоронений как бы указывает на максимум численности и, наоборот. При этом наличие сохранившихся континентальных позднеплейстоценовых отложений, которые также датируются радиоуглеродным методом, определяет доступность захоронений для их нахождения в настоящее время. То есть мы можем однозначно утверждать лишь то, что количество находок захоронений мамонта моложе 10 тыс. лет единично, так как площадь обнажения голоценовых отложений максимальна. Соответственно, сокращение находок древнее 60 тыс. лет может быть связано: а) с ограничениями радиоуглеродного метода датировок (омолаживание); б) с малой сохранностью или сокращением по
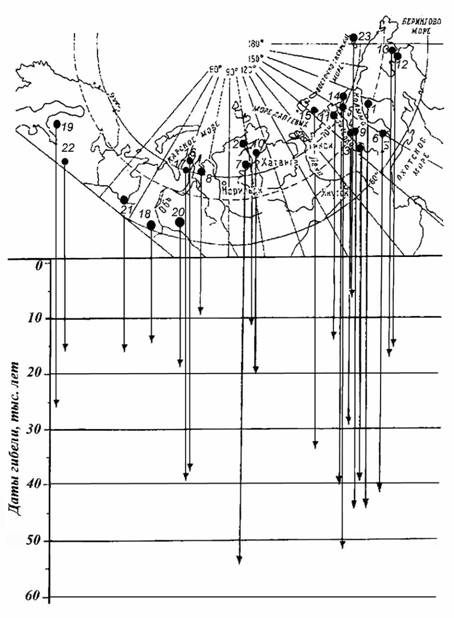
Рис. 1. Некоторые места и хронология находок в Евразии мерзлых туш, скелетов и их фрагментов мамонтовой фауны плейстоцена и голоцена (по Украинцевой В.В. [14], с изменениями и дополнениями):
1 – мамонт Герца (Березовский), 1900; 2 – мамонт р. Мамонта, 1948; 3 – лошадь Черского, 1968; 4 – мылачхинский бизон, р. Индигирка, 1971; 5 – мамонт Русанова (Шандринский), 1971; 6 - киргиляхский мамонт (Дима), 1977; 7 – мамонт Верещагина (Хатангский), 1977; 8 – юрибейский мамонт, 1979; 9 – мамонт р. Тирехтях, 1971; 10 – мамонт Жаркова, р. Большая Балахня, 1997; 11 – мамонты р. Берелех, 1970; 12 – мамонт залива Онемен (Анадырь), 2003, 13 – мамонты р. Канчалан, 1994; 14 - мамонт р. Сундрун (Индигирка), 1971; 15 - Ляховский мамонт (Новосибирские о-ва), 1976; 16 – Ямальский мамонт (Маша), 1988; 17 - Ямальский мамонт (Люба), 2007; 18 – Луговское кладбище мамонтов (Ханты-Мансийский АО), 1950-99; 19 – мамонты Куопио (Финляндия), 1873; 20 – мамонты в Красном Яру (Ханты-Мансийский АО), 2000; 21 – Пермский мамонт, 1927; 22 – мамонт Грязовца (Вологодская область), 2006; 23 – мамонт о-ва Врангеля, 1993
площади выходов на дневную поверхность континентальных отложений среднего и раннего плейстоцена. Уменьшение же количества находок в те или иные периоды времени в интервале 60-10 тыс. лет могут быть следствием не их изначального отсутствия, а эрозией континентальных отложений соответствующего возраста. Иначе говоря, достоверной информации о хронологии вымирания мамонтов не так уж и много.
Ландшафтно-климатические аспекты
Разными авторами отмечалось, что отдельные находки ископаемой фауны приходятся не только на периоды потеплений, но и похолоданий климата и, более того, на максимумы оледенения. Известны, например, находки в Скандинавии, когда гибель и захоронение мамонтов произошли в условиях сплошного щитового ледника [2, 4, 5] Эти особенности, с одной стороны, ставят под сомнение суждения о прямой связи потепления с гибелью животных, с другой указывает на существование более значимого фактора влияющего на количество находок мамонтовой фауны. Таким фактором, по-видимому, являются благоприятные для захоронения и сохранения ископаемых остатков ландшафтные условия. В большинстве случаев это условия перигляциальной зоны ледников с многочисленными реками, ручьями, оврагами, озерами и болотами. Здесь же собственно и накапливаются ледниковые и водно-ледниковые отложения, максимально проявлен криогенез осадков. Погибшие же по тем или иным причинам животные в условиях ледника рано или поздно при его отступлении в виде фрагментов скелета или в состоянии замороженного трупа также оказываются в перигляциальной зоне. Значит, не существует прямой зависимости между климатом и гибелью животных, а ареал обитания мамонта и его плейстоценовых спутников мог распространяться как на перигляциальную зону, так и на ледник – покровный, сетчатый, предгорный, которые вместе и образуют пространство сезонных и многолетних миграций мамонтовой фауны.
О неоднозначности влияния изменений климата на популяцию мамонта свидетельствуют и несложные подсчеты распределения находок ископаемой фауны по климатическим эпохам. На сайте известного исследователя Севера С.А. Зимова приводятся такие данные: «…на тёплые эпохи приходится 48,7 %, на холодные - 30,8 %, на переходные периоды 20,5 % находок. На основании полученных данных, становится очевидным, что популяция мамонтов Сибири вплоть до 12 тыс. лет назад успешно переживала неоднократные колебания климата и изменения растительных зон от межледниковых к ледниковым и наоборот. Объяснить резкое сокращение ареала мамонтов и их последующее вымирание… только общим изменением климата в сторону потепления не удается». Кроме того, общее потепление климата, изменения в растительном покрове и коренные изменения в перигляциальном ландшафте, связанные с коренной перестройкой ландшафта не синхронны во времени. Так по данным А.Н. Котова в начале эпохи голоценового потепления около 14-13 тыс. лет назад на фоне глобального изменения климата на северо-востоке Азии наблюдалось снижение скоростей осадконакопления. Активизация процессов термокарста и термоэрозии по породам ледового комплекса произошла только 10-8 тыс. лет назад, спустя 4 тыс. лет после потепления климата [6]. Позднее мы вернемся к этому важному выводу.
К ландшафтным условиям также имеет отношение характер захоронений. Специфичность этой части проблемы вымирания фауны отмечал палеонтолог и писатель И.А Ефремов, который и предложил научное направление – изучение исследований условий гибели и захоронения ископаемой фауны – тафономию. Не рассматривая пока ископаемые останки мамонта, найденные на стоянках палеолитического человека, выделим три основных их типа. В первом случае это находки практически целых замороженных трупов или их крупных фрагментов, принадлежащих единичным животным. В этом случае, скорее всего мы имеем дело с коренным залеганием объекта в ландшафтных условиях соответствующих его гибели и захоронению. Второй, особенный по нашему мнению тип, связан с массовыми монозахоронениями групп животных, по-видимому, мамонтовых семей или, иначе говоря, матриархальных прайдов. В основном это захоронения костей или частей скелетов, позволяющих их практически полностью реконструировать. Следует полагать, что это перемещенное, дислоцированное или нарушенное захоронение в ландшафтных условиях близких к обстановке захоронения животных, но отличных от него по условиям гибели. И, наконец, третий тип это скопления костей крупных млекопитающих, в том числе хищников, копытных и грызунов. Общие захоронения обнаруживаются в основном в надпойменных террасах крупных рек или эстуариев. Находят захоронения по наиболее сохранившимся прочным костным останкам крупных млекопитающих бивням, черепам, берцовым костям мамонта, носорога, бизона, лошади. Хотя в последующем, в ходе специальных исследований обнаруживаются в той или иной степени сохранившиеся кости других более мелких животных. Можно предположить, что имело место многократное и поэтапное перемещение разновозрастных останков с обширной площади в ходе водной и ледниковой эрозии и аккумуляции. Условия захоронения и сохранность костных останков могут лишь указывать на пределы территории, с которой они мобилизованы, на расстояние перемещения и ландшафтную обстановку последнего захоронения. Таким образом, с точки зрения изучения «гибельных» для мамонтов ландшафтов интерес представляют единичные захоронения замороженных трупов и моно захоронения групп мамонтов. Общим для первого и второго типа находок ископаемой фауны является едома – комплекс континентальных сильнольдистых тонкодисперсных отложений, которая в свою очередь сама является плейстоценовой загадкой [1].
Антропогенный фактор и его роль
Но перейдем к антропогенной составляющей проблемы вымирания мамонта. Именно с ней связана одна из самых популярных и аргументируемых, особенно среди североамериканских ученых, гипотеза исчезновения мамонтовой фауны в позднем плейстоцене - голоцене. В числе основных доводов истребление человеком крупных млекопитающих - единая хронология расселения человека с исчезновением «охотничьих видов» крупных животных, многочисленные находки костных останков на стоянках человека мезолита и палеолита, охотничий азарт человека охотника, когда уничтожается больше животных, чем необходимо для потребления [7-9]. Есть и другие аргументы, которые зачастую идеологизированы и проводят аналогию с пагубным влиянием современной цивилизации на животный мир.
Совпадение по времени расселения человека по северу Евразии и Северной Америке, заселение Индонезийского архипелага и Австралии с коренной перестройкой фауны этих земель в сторону исчезновения крупных животных, действительно веский аргумент. В Австралии, например, вымирали крупные животные, не принадлежащие к мамонтовой фауне. Однако на северо-востоке Евразии и на Аляске до сих пор не обнаружено столь большого числа стоянок палеолитического человека охотника, а найденные, свидетельствуют, что основной их добычей были северные олени, бизоны и другие, более мелкие млекопитающие [10]. Да и в Австралии совместное «безоблачное» проживание заселившегося человека и местной фауны продолжалось несколько тысяч лет [4] Более осторожные сторонники антропогенной гипотезы вымирания мамонтов предполагают, что первобытный человек, лишь завершил охотой исчезновение плейстоценовых животных, популяции которых были подорваны влиянием голоценового потепления. Но даже в этом случае трудно представить себе группы охотников «гурманов» в поисках последних мамонтовых прайдов для истребления, в то время когда рядом в избытке более доступная добыча – северные олени, бизоны, лоси [11]. Так же трудно поддается реконструкции технологии транспортировки добытой туши мамонта к стойбищу или стоянке древнего человека. Представляется, что в позднем плейстоцене – голоцене и ранее мамонт был случайной добычей попавших в природную ловушку, недавно умерших животных или представлял собой предмет опасной ритуальной охоты. Так же как для древних морских зверобоев - гренландский, горбатый, серый киты - редкая и ритуальная добыча против обычного промысла моржей и тюленей.
Эпоха активного использования человеком костных останков мамонта в качестве материала для строительства жилища и топлива приходится на период расцвета популяции животных в западной части Евразии. Территория размещения стоянок, охоты и рыбалки древнего человека была приурочена к перигляциальной зоне и изменялась с перемещением границ ледников [12]. Именно в перигляциальной зоне находятся как монозахоронения мамонтов, так и общие захоронения останков мамонтовой фауны. Стоит предположить, что такие захоронения оттаивали и в древности, а в условиях дефицита древесного материала и орудий для его обработки служили для строительства и отопления жилища. Это конечно не исключает охоту на мамонта палеолитических охотников и утилизацию отходов потребления в обустройстве жилища, но доля полученного таким образом материала вряд ли была значительна.
Завершая анализ роли древнего человека в исчезновении мамонта, невольно испытываешь желание предложить свою оригинальную гипотезу. Например, связывающую перестройку фаунистического комплекса в конце плейстоцена с воздействием антропогенного фактора, но не в качестве причины вымирания мамонтов, а как необходимое условие сохранения отдельных представителей мамонтовой фауны – лошади, быка, верблюда, северного оленя. Древний человек охотник, кочевавший вслед за стадами диких животных по обширным просторам перигляциальной зоны, безусловно, с усложнением социального устройства и уменьшением численности объектов охоты, занялся доместикацией животных мамонтового комплекса и делал первые шаги к животноводству. Недостаток археологических находок этого времени легко объясняется кочевым образом жизни «протоскотоводов». Все одомашненные животные из числа мамонтовой фауны сохранились и в настоящее время, если не в диком, то в домашнем виде. Причем популяции диких и домашних животных, например, северных оленей на Чукотке, оказываются связанными между собой не только пространственно, но и генетически. Одомашненные животные не были так зависимы от флуктуаций климата и перестройки природной среды. Направляемые разумным человеком стада не были так жестко привязаны к традиционным путям миграции диких животных и перемещались к сохранившимся в плейстоцене-голоцене или вновь образовавшимся пастбищам. В последующем, домашние олени подпитывали численность популяции диких животных или становились таковыми при одичании. Это явление не редкость и в настоящее время, не только на крайнем северо-востоке Евразии, но и на других континентах. Выходит, что шерстистый мамонт позднего плейстоцена вымер скорее не в результате истребления человеком, а потому, что по тем или иным причинам не был им одомашнен.
Вымирание мамонтов и эволюция биосферы
Вслед за эволюционистами логичней допустить, что расселение человека по континентам и перестройка фаунистического и флористического комплекса плейстоцена есть неразделимый и необратимый процесс, обусловленный едиными закономерностями эволюции [10, 13]. В данном ключе перспективен поиск сходств и отличий условий позднеплейстоценового витка эволюции от предшествующих эпох и современности. Несомненный интерес в этом плане представляет мысль об определяющем влиянии на развитие плейстоценовой фауны и флоры: а) резкого усиления зональности природной среды с сокращением ареала интразональных ландшафтов – пойменных и суходольных лугов и лесов; б) нарушение путей сезонной и многолетней миграции плейстоценовых животных, в первую очередь наиболее уязвимых к изменению рациона крупных млекопитающих; в) смешение ресурсно-конкурентных зооценозов, которое приводило к деградации пастбищ и нарушению трофических цепей [3, 14]. Такие изменения могли произойти в исторически короткий промежуток времени, затрудняющий адаптацию к ним зооценозов. О роли миграций в сохранении популяции шерстистого мамонта на фоне несинхронных потеплений Евразии в позднем плейстоцене известно [15]. Ежегодные сезонные миграции перелетных птиц, гнездующихся в Арктике и Субарктике, косвенно подтверждают существования в плейстоцене сложной и масштабной миграционной структуры. В современное время можно наблюдать сезонные и многолетние цикличные миграции дикого северного оленя с юга на север и с запада на восток, миграции лосей вслед за тающими снежниками, за молодой порослью ивы и нивальными луговинами.
Количество гипотез объясняющих конкретные причины вымирания мамонтов, с появлением новых находок ископаемой фауны и совершенствованием аналитических методов, будет расти и дальше. Но все они, так или иначе, будут укладываться в формулу общих закономерностей эволюции биосферы Земли – геологической, климатической, ландшафтной, ценотической, антропогенной. Оттого загадка вымирания мамонтов обладает важным палеогеографическим и палеоландшафтным содержанием, которое сокрыто в реконструкции условий гибели и захоронения животных. Именно эта сторона проблемы актуальна сегодня, когда все более востребованы модели изменения биосферы Земли в условиях глобального потепления или похолодания климата. Рассмотрим еще несколько оригинальных гипотез, раскрывающих общности и особенности ландшафтных обстановок и климатических условий обитания мамонтовой фауны на рубеже плейстоцена - голоцена, которые могут быть полезными для прогнозирования характера и последствий ожидаемого глобальных изменений среды в нынешнее время.
Лёссы как среда обитания мамонтовой фауны
Очевидный интерес представляет отмеченный нами выше факт, что сезонными и многолетними миграциями мамонтов была охвачена не только перигляциальная зона, но и ледники в различных их проявлениях. Чем же питались на обширных ледяных просторах столь крупные животные? Этот вопрос неразрывно связано с другим – происхождением лёссов, широко развитых в Евразии и Северной Америке [2]. Не вдаваясь в подробности аргументаций водной и эоловой гипотез, отметим общую закономерность современного распространения лёссовидных отложений. Они практически отсутствуют в областях покровного и горно-долинного оледенений, которые пространственно вклиниваются в перигляциальную зону, образуя с ней сложную извилистую границу. На картах лёссовых пород бывшего СССР они имеют сплошное распространение к югу от 60° широты и до 90° долготы. Далее на восток и северо-восток лёссы покрывают шлейфом увалистые равнины и мелкосопочники Забайкалья, рек Амур, Енисей, Лена, Индигирка, Колыма, Анадырь, Пенжина, Паляваам. Тем самым, можно предположить, что области оледенений в эпохи межледниковья, в том числе на рубеже плейстоцена и голоцена являлись областями водного сноса лёссового материала изначально накопленного эоловой аккумуляцией. Косвенно эоловую активность эпохи плейстоцена подтверждают результаты бурения льдов Гренландии и Антарктиды, содержание в которых твердых примесей 10 тыс. лет назад в десятки и сотни раз превышает современный уровень. Особое положение занимают лёссовидные криогенные отложения северо-востока Евразии, где эоловому и водному их накоплению сопутствовали процессы подземного льдообразования [10, 16, 17]. Главный же вывод в том, что в ксерофитные эпохи засушливого, контрастного климата обширные пространства северного полушария, как занятые ледниками, так и свободные от них были покрыты шлейфом тонкодисперсных слабо выщелоченных осадков благоприятных для развития холодоустойчивых, закустаренных тундростепей [13]. Оторфованные супеси и супесчаные суглинки с плотным и высоким травостоем, обеспеченные влагой, подстилающего их льда и многолетнемерзлых пород являлись прекрасными тундростепными пастбищами и теплоизоляторами подземного жильного и поверхностного покровного льда. Возможно, также, что именно эта теплоизоляция является причиной запаздывания термокарста и термоэрозии в голоцене по отношению к началу глобального потепления климата [6]. Выходит, что в плейстоцене мамонты, а вместе с ними многочисленные стада травоядных животных в ходе миграций свободно перемещались по перигляциальной зоне, предгорным и покровным, а возможно и горно-долинным ледникам невысоких горных массивов [3].
Любопытно, что еще в начале прошлого века было распространено мнение, что сильнольдистые лёссовидные отложения побережья и островов Восточно-Сибирского и Чукотского морей представляют собой захороненные покровные ледники. Сходство для неискушенного натуралиста может быть поразительным (рис. 2).

Рис. 2. Береговой обрыв р. Майн с обнажением повторно-жильного льда в едомной толще
В последующем это гипотеза была опровергнута моделью сингенетического образования повторно-жильных льдов, но не утратила своей привлекательности для других территорий и исторических эпох.
Термокарстовая катастрофа
Потепление климата, в первую очередь проявлялось в увеличении сезонно-талого слоя и активизации термокарста и термоэрозии по перекрытым лёссом поверхностным и подземным жильным и пластовым льдам [4]. Результатом этих процессов стало образование многочисленных глубоких «подземных» ледяных гротов и лабиринтов пещер, перекрытых сверху тонким слоем льда, лёссовых отложений, почв и кустарниково-травяной дерниной.
Таким образом, в период потепления климата ледниковые пастбища мамонтовой фауны оказались усеяны ловушками холодильниками, специализированными на крупных животных, передвигающихся группами одними и теми же маршрутами. Во многом сходная обстановка складывалась в перигляциальной зоне горно-долинных ледников и в Берингийских тундростепях, где вытаивали мощные подземные сингенетические повторно-жильные льды. Следовательно, обильные тундростепные пастбища не только постепенно деградировали с потеплением климата, разрушались с таянием ледников, но и стали недоступны мамонтам и другим травоядным животным по причине банальной изоляции их ледяными разломами. Мамонтовая фауна больше не могла беспрепятственно мигрировать по необъятным просторам тундростепей, а доступные пастбища просто вытаптывались ими в замкнутом термокарстовыми провалами пространстве.
Впервые гипотезу о захоронении мамонтов в термокарстовых провалах – ледяных ловушках едомы в тундростепях северо-востока Евразии высказали О.Ф. Гольц в 1901 и М.И. Папернов, Ш.Ш. Гасанов в 1961 году [1, 11] Однако явление это не было оценено ни по своему масштабу, ни по значимости. Подземные термокарст и термоэрозия протекают и в настоящее время, хотя и при меньшем размахе. В связи с чем, необходимо отметить две важные их особенности: а) формирование термокарстовых провалов отличает скоротечность и катастрофический характер - образование их происходит на большой протяженности и в короткий срок; б) за образованием провала следует обрушение кровли и оплывание бортов, которые закрывают и теплоизолируют его днище. Это и объясняет плейстоценовые находки хорошо сохранившихся замороженных трупов мамонтов и диких лошадей.
Дальнейшая судьба захоронений связана с природой термокарстового провала – ловушки холодильника. Термокарстовые провалы по сингенетическим повторно-жильным льдам могут быть на века «запечатаны» в многолетнемерзлой толще, а в последующем размыты с перезахоронением костных останков. В условиях нестационарного предгорного ледника провалы могут многократно закрываться и образовываться вновь, а костные останки надолго сохраняться и перемещаться вместе с ледниковыми отложениями. Это может быть одной из причин моно захоронений мамонтов и крупных скоплений костных останков плейстоценовой фауны в одном месте. Важно отметить, что в роли ловушек холодильников могли выступать не только термокарстовые провалы межледниковий, но и ледяные расщелины в ледниковые периоды, хотя, наверное, и в меньших масштабах.
Очевидно, что массовый термокарст и термоэрозия были присущи всем межледниковым эпохам. Косвенное подтверждение тому плейстоценовые бедленды – реликтовые гляциальные (термокарстовые) ландшафты равнин Азии и Северной Америки, ошибочно, по нашему мнению, интерпретируемые как результат залпового сброса ледниковых озер [18]. Рассматривая явление в таком масштабе можно найти аналогию с последствиями всемирного потопа, ибо результат экзогенно-криогенных процессов это не только изменение форм рельефа поверхности, но и образование специфичных отложений с большой долей пылевато-илистого материала в форме потоков и разнообразных оплывин, напоминающих отложения ливневых стоков.
Остается открытым все тот же вопрос, почему именно голоценовое потепление завершило эпоху мамонтов. Здесь стоит вернуться к уже описанным выше закономерностям эволюции природной среды. А именно усиление климатической и природной зональности, сокращение ареала интразональных ландшафтов, которые, несомненно, играли большую роль в миграции мамонтовой фауны. При этом накануне, в конце плейстоцена на северо-востоке Евразии и в Северной Америке достигли своего расцвета тундростепные ландшафты, которые в некотором роде также были интразональными. Максимальная их биомасса и биопродуктивность способствовали не только росту численности травоядных животных, но и возросшей в конце плейстоцена теплоизоляции льдов и многолетнемерзлых пород от влияния атмосферного потепления. Как уже указывалось выше с опозданием к потеплению климата, но тем катастрофичней и масштабней на рубеже голоцена проявились термокарст и термоэрозия как по ледникам, так и по сингенетическим повторно-жильным льдам берингийских тундростепей. Единичные популяции мамонтов нашли прибежище на изолированной суше с преимущественно скальным основанием, подобно острову Врангеля и просуществовали там, в карликовых формах, до начала современной эпохи [5].
Заключение
Приблизительно такой сценарий событий прослеживается нами на рубеже плейстоцена и голоцена. Голоценовое потепление с одной стороны является завершением очередного ледникового цикла, с другой очередным этапом - витком эволюции биосферы антропогена, выразившимся в усложнении структуры и контрастности природной среды. Исчезновение, а точнее глубокое преобразование мамонтовой фауны и флоры как составной части эволюции биосферы было континуально-дискретным или закономерно-случайным событием. В роли фактора случайности, дискретности эволюции выступила массовая и масштабная термокарстовая катастрофа, хотя на ее месте могли бы быть всемирный потоп, падение крупного астероида, вулканическая активность Земли.
Мое знакомство с мамонтовой проблемой ограничивалось случайными находками ископаемой фауной в ходе изучения современных ландшафтообразующих процессов в Чукотской тундре. В основном это смешанные костные останки в водно-ледниковых отложениях позднего плейстоцена перигляциальной зоны. В 2005 году в ходе мониторинга термоабразии береговых обрывов залива Онемен эстуария Анадырь. Это был хорошо сохранившейся зуб небольшого мамонта, обугленные кости лошади и мелких млекопитающих (рис. 3).

Рис. 3. Первые находки плейстоценовой фауны в эстуарии Анадырь
Через год, здесь же в обрыве обнажились целый бивень и крупный обломок бивня хорошей сохранности длиной 175 и 86 см. Вмещают костные останки оторфованные супесчано-илистые сильнольдистые многолетнемерзлые породы, которые подстилают современные тундровые торфянистые почвы. Ниже по разрезу залегает толща ледниково-морских слоистых глинисто-песчаных отложений, датированных радиоуглеродным методом в интервале 32-41 тыс. лет. Самая молодая известная радиоуглеродная датировка растительных остатков обрывов залива Онемен 12790 лет. Эти факты позволили определить данное захоронение как дальнопереносное, несвязанное с коренным обитанием мамонов образованное в палеоустье реки Анадырь.
[1] В геоморфологии термином едома означают небольшую плосковершинная возвышенность, холм до десятков метров относительной высоты, сложенную сильнольдистыми четвертичными отложениями.
[2] Известно около 20 гипотез происхождения лёссов, объясняющих их генезис накоплением эоловых, аллювиально-озерных, склоновых осадков, почвообразованием и мерзлотными процессами.
[3] На северо-востоке Евразии существенная часть находок мамонтовой фауны, в том числе мамонтенка Димы приходится на предгорье в зоне плейстоценового горно-долинного оледенения.
[4] Здесь следует подчеркнуть, что причиной активизации термокарста и термоэрозии может быть не только увеличение среднегодовой температуры воздуха атмосферы, но гумидизация климата, например, в условиях дождливого лета, мягкой и снежной зимы.
[5] В средневековых арабских записях упоминается об охоте конных монголов на территории нынешнего Забайкалья на волосатого единорога, как считал академик И.Г. Подопличко – шерстистого носорога [4]
Библиография
1. Верещагин Н.К. Почему вымерли мамонты. – Л.: Наука, 1979. 195 с.
2. Васильчук Ю.К., Васильчук А.К., Лонг О. и др. Новые данные о популяции мамонтов в позднеплейстоценовой криолитозоне Евразии // Доклады академии наук, том 360, № 6, 2000. С. 815-816
3. Тихонов А.Н. Мамонт. (Серия «Разнообразие животных» Вып. 3). Москва – С.-Петербург: Т-во научных изданий КМК. 2005. 90 с.
4. Никонов А. «Россия – Родина слонов»? Нет мамонтов // ЗНАНИЕ-СИЛА, № 11, 2002. С. 60-67.
5. Donner J., Jungner H., Kurten B. // Bull/Geol. Soc. Finland. V. 5. N 1/2, 1979. P. 45-54
6. Котов А.Н. Плейстоценовые оледенения южной части Нижнеанадырской впадины и севера Корякского нагорья // Четвертичная палеогеография Берингии. Магадан: МПО СВНЦ ДВО РАН, 2002. С.67-75.
7. Аникович М.В. Палеолитическая Мекка // ЗНАНИЕ-СИЛА, № 1, 1996. с. 60-70
8. Аникович М.В. Повседневная жизнь охотника на мамонтов. – М.: «Молодая гвардия». 2004, 314 с.
9. Шумилов Ю. Старое и новое в мамонтовой судьбе // Наука и Жизнь, № 7, 2004.
10. Шило Н.А. Четыре космопланетарных проблемы: от Солнечной системы до Каспия. – М.: Фонд «Новое тысячелетие», 1997. 136 с.
11. Русанов Б.С. Внимание: мамонты! – Магаданское книжное издательство, 1976. 192 с.
12. Мащенко Е.Н. Последние мамонты Русской равнины // Химия и Жизнь. № 8, 2000. С. 32-37.
13. Трегубов О.Д. Об устойчивости тундр к техногенному воздействию и глобальным изме-нениям среды // Вестник ДВО РАН. № 4, 2010. С. 79-89
14. Украинцева В.В. Растительность и климат Сибири эпохи мамонта. – Красноярск: Восточ-но-Сибирский филиал Международного института леса. 2002. 192 с.
15. Шер А.В. Природная перестройка в Восочно-Сибирской Арктике на рубеже плейстоцена и голоцена и ее роль в вымирании млекопитающих и становлении современных экосистем (сообщение 1) // Криосфера Земли, т. 1, № 1. 1997. С.21-30
16. Томирдиаро С.В. Вечная мерзлота и освоение горных стран и низменностей. Магадан-ское книжное издательство, 1972. 174 с.
17. Васильчук Ю. К., Васильчук А.К. Повторно-жильные льды долины реки Майн и реконструкция зимних палеотемператур воздуха южной Чукотки 38-12 тысяч лет назад // Криосфера Земли, т. XXI, № 5. 2017. С. 27-41.
18. Силкин Б. Пол-Азии под Водой // ЗНАНИЕ-СИЛА, № 9, 2003. 66 с.
References
1. Vereshchagin N.K. Pochemu vymerli mamonty. – L.: Nauka, 1979. 195 s.
2. Vasil'chuk Yu.K., Vasil'chuk A.K., Long O. i dr. Novye dannye o populyatsii mamontov v pozdnepleistotsenovoi kriolitozone Evrazii // Doklady akademii nauk, tom 360, № 6, 2000. S. 815-816
3. Tikhonov A.N. Mamont. (Seriya «Raznoobrazie zhivotnykh» Vyp. 3). Moskva – S.-Peterburg: T-vo nauchnykh izdanii KMK. 2005. 90 s.
4. Nikonov A. «Rossiya – Rodina slonov»? Net mamontov // ZNANIE-SILA, № 11, 2002. S. 60-67.
5. Donner J., Jungner H., Kurten B. // Bull/Geol. Soc. Finland. V. 5. N 1/2, 1979. P. 45-54
6. Kotov A.N. Pleistotsenovye oledeneniya yuzhnoi chasti Nizhneanadyrskoi vpadiny i severa Koryakskogo nagor'ya // Chetvertichnaya paleogeografiya Beringii. Magadan: MPO SVNTs DVO RAN, 2002. S.67-75.
7. Anikovich M.V. Paleoliticheskaya Mekka // ZNANIE-SILA, № 1, 1996. s. 60-70
8. Anikovich M.V. Povsednevnaya zhizn' okhotnika na mamontov. – M.: «Molodaya gvardiya». 2004, 314 s.
9. Shumilov Yu. Staroe i novoe v mamontovoi sud'be // Nauka i Zhizn', № 7, 2004.
10. Shilo N.A. Chetyre kosmoplanetarnykh problemy: ot Solnechnoi sistemy do Kaspiya. – M.: Fond «Novoe tysyacheletie», 1997. 136 s.
11. Rusanov B.S. Vnimanie: mamonty! – Magadanskoe knizhnoe izdatel'stvo, 1976. 192 s.
12. Mashchenko E.N. Poslednie mamonty Russkoi ravniny // Khimiya i Zhizn'. № 8, 2000. S. 32-37.
13. Tregubov O.D. Ob ustoichivosti tundr k tekhnogennomu vozdeistviyu i global'nym izme-neniyam sredy // Vestnik DVO RAN. № 4, 2010. S. 79-89
14. Ukraintseva V.V. Rastitel'nost' i klimat Sibiri epokhi mamonta. – Krasnoyarsk: Vostoch-no-Sibirskii filial Mezhdunarodnogo instituta lesa. 2002. 192 s.
15. Sher A.V. Prirodnaya perestroika v Vosochno-Sibirskoi Arktike na rubezhe pleistotsena i golotsena i ee rol' v vymiranii mlekopitayushchikh i stanovlenii sovremennykh ekosistem (soobshchenie 1) // Kriosfera Zemli, t. 1, № 1. 1997. S.21-30
16. Tomirdiaro S.V. Vechnaya merzlota i osvoenie gornykh stran i nizmennostei. Magadan-skoe knizhnoe izdatel'stvo, 1972. 174 s.
17. Vasil'chuk Yu. K., Vasil'chuk A.K. Povtorno-zhil'nye l'dy doliny reki Main i rekonstruktsiya zimnikh paleotemperatur vozdukha yuzhnoi Chukotki 38-12 tysyach let nazad // Kriosfera Zemli, t. XXI, № 5. 2017. S. 27-41.
18. Silkin B. Pol-Azii pod Vodoi // ZNANIE-SILA, № 9, 2003. 66 s.
Ссылка на эту статью
Просто выделите и скопируйте ссылку на эту статью в буфер обмена. Вы можете также
попробовать найти похожие
статьи
|
|



