|
ГЛАВНАЯ
> Вернуться к содержанию
Физика биологии и медицины
Правильная ссылка на статью:
Бинги В.Н., Рубин А.Б.
О квантовой природе магнитных явлений в биологии
// Физика биологии и медицины.
2023. № 1.
С. 44-73.
DOI: 10.7256/2730-0560.2023.1.40435 EDN: SVLAQR URL: https://nbpublish.com/library_read_article.php?id=40435
О квантовой природе магнитных явлений в биологии
Бинги Владимир Николаевич
ORCID: 0000-0003-1341-9591
доктор физико-математических наук
ведущий научный сотрудник, Институт общей физики им. А.М. Прохорова РАН
119991, Россия, г. Москва, ул. Вавилова, 38
Binhi Vladimir
Doctor of Physics and Mathematics
Head of Laboratory, Prokhorov General Physics Institute of the Russian Academy of Sciences
38 Vavilova str., Moscow, 119991, Russia

|
vnbin@mail.ru
|
|
 |
Другие публикации этого автора
|
|
|
Рубин Андрей Борисович
доктор биологических наук
заведующий кафедрой, академик РАН, Московский государственный университет им. М.В. Ломоносова
119234, Россия, г. Москва, ул. Ленинские Горы, 1/12
Rubin Andrei Borisovich
Doctor of Biology
Head of Department, Lomonosov Moscow State University, Academician of the RAS
119234, Russia, Moscow, Leninskie Gory str., 1/12
|
|
rubin@biophys.msu.ru
|
|
 |
|
DOI: 10.7256/2730-0560.2023.1.40435
EDN: SVLAQR
Дата направления статьи в редакцию:
12-04-2023
Дата публикации:
08-06-2023
Аннотация:
В обзоре обсуждаются микроскопические механизмы действия слабых магнитных полей на организмы. Магнитобиология различает магниторецепцию – действие магнитного поля на специализированные рецепторы – и неспецифический отклик, развивающийся в отсутствие таких рецепторов. Неспецифические эффекты слабых магнитных полей обладают общностью и универсальностью: они имеют место во всех организмах. Часто эти эффекты маскируются под результат действия неконтролируемых случайных факторов, проявляются в виде увеличенного разброса измерений и сопровождаются малой воспроизводимостью. Рассмотрены проблемы объяснения биологических эффектов слабых магнитных полей, сравнимых по величине с геомагнитным полем. Определены наиболее правдоподобные механизмы. Показано, что природа магнитного действия связана с квантовой динамикой магнитных моментов электронов, магнитных ядер и, возможно, вращений молекулярных групп. В числе наиболее обоснованных, прежде всего, спин-химический механизм. Его известная низкая чувствительность к слабым магнитным полям может быть усилена на два-три порядка включением спин-коррелированных радикальных пар в работу биополимерных ферментов, в частности, рибосомальных. Показано, что исследование действия значительно ослабленных магнитных полей, – в сравнении с геомагнитным, – на клеточные процессы имеет перспективы разнообразных практических применений. Перечислены механизмы, которые предлагались для объяснения неспецифических эффектов, но оказались несостоятельны.
Ключевые слова:
квантовый эффект, магнитобиология, слабое магнитное поле, гипомагнитное поле, неспецифический эффект, случайный эффект, молекулярный механизм, проблема кТ, спиновая химия, механизм радикальных пар
Abstract: The review discusses the microscopic mechanisms of the action of weak magnetic fields on organisms. Magnetobiology distinguishes between magnetoreception, i.e., the effect of a magnetic field on specialized receptors, and a nonspecific response that develops without such receptors. The nonspecific effects of weak magnetic fields are highly general and universal: they occur in all organisms. Often these effects are disguised as the result of the action of uncontrolled random factors, appear as an increased scatter of measurements, and accompanied by low reproducibility. The nature of nonspecific magnetic effects, as is shown in this review, is related to the quantum dynamics of the magnetic moments of electrons, magnetic nuclei, and, possibly, rotations of molecular groups. Among the most substantiated is the spin-chemical mechanism, first of all. Its known low sensitivity to weak magnetic fields can be increased by including spin-correlated radical pairs in the enzymes that catalyze biopolymer processes, e.g., ribosomal ones. We show that research on the effects of significantly weakened magnetic fields compared with the geomagnetic field on cellular processes has prospects for various practical applications. The mechanisms proposed to explain nonspecific effects, but turned out to be untenable, are listed.
Keywords: quantum effect, magnetobiology, weak magnetic field, hypomagnetic field, nonspecific effect, random effect, molecular mechanism, kT problem, spin chemistry, radical pair mechanism
Введение
Пионером научного изучения биологических эффектов магнитного поля (МП) является, по-видимому, наш соотечественник акад. А.Ф. фон Миддендорф (1815–1894, Российская империя). Основываясь на своих наблюдениях за перелетными птицами, он писал: «Внутреннее магнитное чувство наших ‘парусников неба’ берет на себя функцию, выполняемую компасом на кораблях» [1] с. 9, рис. 1.

Рис. 1. Титульная страница монографии А.Ф. фон Миддендорфа с материалами экспедиции в Сибирь.
Практическое изучение биологических эффектов МП изначально связано с медициной. Известно, например, что уже в 1750-х годах немецкие врачи изучали терапевтический потенциал МП [2]. В литературе имеются свидетельства того, что в XIX и первой половине XX веков десятки врачей, в том числе отечественные, С.П. Боткин, А.В. Селезнев, В.И. Кармилов и др., пробовали использовать постоянные магниты и искусственные постоянные и переменные МП в физиотерапевтических целях [3,4]. Однако зарождение исследований именно природы явления было связано с разработкой в СССР и США в 1960-х годах излучателей электромагнитных волн миллиметрового диапазона. Оказалось, что микроволны способны действовать на растения и микроорганизмы [5–7], что подтверждено многочисленными современными исследованиями, см. напр. [8–10]. Это выглядело парадоксальным, так как энергия квантов такого поля была на два порядка меньше энергии активации химических превращений, а частотная избирательность действия и малая мощность излучения исключали тепловую природу эффекта.
В развитие этих исследований уже в 1970-х и 1980-х годах наблюдали биологические эффекты слабых низкочастотных МП, преимущественно ниже уровня напряженности геомагнитного поля (геоМП) и в частотном диапазоне от единиц до сотен Гц [3,11]. Возник вопрос: влияют ли фоновые МП, сопровождающие передачу и использование электроэнергии, на здоровье? В 1983 г. для координации исследований природы этого явления и разработки терапевтических методов была создана Проблемная комиссия Минздрава СССР «Магнитобиология и магнитотерапия в медицине» [12]. В 1992 году Конгресс США утвердил пятилетнюю исследовательскую программу — Исследование электрических и магнитных полей и распространение общественной информации — стоимостью более 40 млн. долларов [13]. Отметим, что к моменту завершения программы в 1998 г. вопрос о влиянии МП на здоровье человека не был решен однозначно. Только последующие десятилетия коллективных усилий исследователей всего мира позволили установить примерную степень опасности фоновых МП [14] стр. 332, [15–20]. В настоящее время достоверно установлено, что МП на порядки меньшие, чем геоМП, могут вызывать биологические эффекты [21–26]. О влиянии слабых МП на организмы опубликовано более десяти тысяч научных работ. Тем не менее такое действие продолжает вызывать вопросы.
Сегодня магнитобиология различает специфические — за счет особых рецепторов — и неспецифические магнитные эффекты [27,28]. Специфические эффекты возникают благодаря специализированным магнитным рецепторам, развитым в ходе биологической эволюции. Они обеспечивают выживание некоторых видов животных при длительных сезонных миграциях. Имеются десятки экспериментов по навигации животных, где в больших вольерах или бассейнах симулировали магнитные условия, характерные для мест сезонных пребываний, разнесенных на несколько сотен километров. Животные при этом кардинально меняли направление движения [29]. В других наблюдениях, магнитные бури меняли направление миграций [30], и т.д. Факты чувствительности некоторых организмов к вариациям МП порядка геомагнитных вариаций надежно установлены.
Специфический магнитный эффект — это магнитное чувство, т.е. магниторецепция [21,31]. Например, у некоторых перелетных птиц есть специальный рецептор, объединенный со зрительной системой, рис. 2. Птица может «видеть» изменения МП порядка десятков нТл и использовать это для ориентации и навигации по магнитному рельефу Земли [32]. Некоторые мигрирующие насекомые обладают аналогичной способностью, также связанной со зрением [33].

Рис. 2. Зарянка Erithacus rubecula, использующая, как полагают, квантовый магнетохимический компас для ориентации и навигации в геоМП во время сезонных перелетов на расстояния до 2000 км. Фото © F.C. Franklin / CC-BY-SA-3.0.
Помимо эффектов магнитной навигации, вариации МП порядка 1 мкТл оказывают влияние на организмы, не обладающие специальными рецепторами. Это и есть неспецифические магнитные эффекты, вызывающие все больший интерес. Неспецифические эффекты не являются магниторецепцией, поскольку таковая предполагает наличие рецепторов. Еще 30 лет назад МП считали фактором, действующим на человека в обход органов чувств [4], то есть в обход специализированных рецепторов. Неспецифические эффекты наблюдаются у многих организмов, от простейших и грибов до насекомых, растений [34,35], рис. 3, рыб, животных и человека [36]. Интерес к этим эффектам возрастает, так как МП могут изменять множество свойств. В частности, изменяется экспрессия генов [37,38]. Другими словами, МП может быть одним из факторов управления синтезом белка. Однако использовать всю мощь этого способа управления генами пока не удается, так как природа первичного биофизического сенсора МП, не являющегося магниторецептором, в организме пока не объяснена. Не выявлен такой сенсор и экспериментально.

Рис. 3. Кресс-салат Arabidopsis thaliana не имеет специальных магнитных рецепторов, но свойства его проростков меняются в слабом МП. Фотографию любезно предоставил проф. А. Батшауэр (Alfred Batschauer).
С 1980 г. по настоящее время для объяснения биологического действия слабых МП было предложено множество физических механизмов и математических моделей. В список можно было бы включить те, которые учитывают: биогенный магнетит и загрязнение магнитными наночастицами; тепловой нагрев и вихревые токи; магнитогидродинамические эффекты и химическую кинетику с бифуркациями; циклотронный, параметрический и стохастический резонансы; фазовые переходы в магнитных полях и эффекты спиновой химии; интерференцию квантовых состояний ионов и молекулярных групп; когерентные возбуждения; метастабильные состояния воды и др. [22]. На сегодняшний день сохранились и получили развитие лишь несколько подходов, но и они остаются гипотезами.
За последние десятилетия было опубликовано несколько обзоров теоретических работ в области магниторецепции [32,39]. Однако обзоров теоретических концепций, направленных на объяснение неспецифических эффектов, не проводилось. Заполнить этот пробел — цель настоящей работы.
В следующем разделе обсуждаются проблемы, возникающие при объяснении биологических эффектов слабых МП. За ним следует раздел, в котором обосновано, что правдоподобные механизмы магнитного отклика организмов учитывают квантовую динамику первичных акцепторов МП — магнитных моментов электронов, в первую очередь. Перечислены возможные квантовые механизмы и объяснена суть спин-химического механизма радикальных пар (РПМ) — механизма, наиболее представленного в научной литературе. Специальные разделы отведены для более подробного обсуждения трудностей этого механизма, обусловленных малым временем тепловой спиновой релаксации, и обсуждения механизма статистического усиления слабых первичных сигналов МП в процессе клеточной трансляции, — механизма, который, возможно, снимает трудности РПМ. Далее следует раздел, в котором показано, что экспериментальное определение механизма биологического действия слабого МП наиболее целесообразно проводить с использованием гипомагнитного поля. Итоги обзора сконцентрированы в Заключении. Приложения 1 и 2 знакомят с квантовыми механизмами, альтернативными РПМ; Приложение 3 содержит описания часто предлагаемых, но несостоятельных механизмов биологического действия слабых МП; объяснено, почему они являются несостоятельными.
Обзор адресован читателю, еще не знакомому с данной областью исследований. Соответственно, стиль изложения материала упрощенный. Это позволяет иллюстрировать сложные квантовые явления в терминах классической физики. При этом неизбежно ограниченный объем обзора может дать лишь общее представление о магнитных биологических явлениях. Более глубокое их понимание потребует обращения к специализированным источникам из списка литературы.
Проблемы объяснения биологических эффектов слабых МП
Так же как электрическое поле взаимодействует с электрическими зарядами, МП взаимодействует с магнитными моментами. На этом основан любой молекулярный механизм магнитных эффектов в организмах. Магнитный момент — это физическая величина, описывающая магнитные свойства объекта. Известно, например, что электрон имеет как механический момент, или спин, так и магнитный, коллинеарный спину. Классическая аналогия — это волчок, несущий заряды на своем ободке. Круговое движение зарядов создает МП, направленное по оси волчка, — это, по сути, МП микроскопического магнита. Его величина и направление задаются вектором магнитного момента 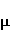 . .
Во внешнем МП 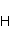 магнитный момент ведет себя подобно стрелке компаса. Внешнее МП старается развернуть момент так, чтобы их направления совпали, т.е. чтобы энергия их взаимодействия магнитный момент ведет себя подобно стрелке компаса. Внешнее МП старается развернуть момент так, чтобы их направления совпали, т.е. чтобы энергия их взаимодействия 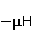 (система СГС) была минимальной. Этому однако препятствует закон сохранения механического углового момента, жестко связанного с магнитным. В результате возникает т.н. прецессия магнитного момента в МП, аналогичная прецессии волчка в поле силы тяжести. Это и вызывает, в конечном счете, биологический отклик организма на МП. В терминах квантовой механики МП, действуя на квантовое состояние магнитных моментов электронов, меняет их спиновое состояние, которое, как известно, является фундаментальным химическим фактором. Сразу же отметим, что процессы с участием спина — это квантовые процессы, описание которых на языке классической физики невозможно. (система СГС) была минимальной. Этому однако препятствует закон сохранения механического углового момента, жестко связанного с магнитным. В результате возникает т.н. прецессия магнитного момента в МП, аналогичная прецессии волчка в поле силы тяжести. Это и вызывает, в конечном счете, биологический отклик организма на МП. В терминах квантовой механики МП, действуя на квантовое состояние магнитных моментов электронов, меняет их спиновое состояние, которое, как известно, является фундаментальным химическим фактором. Сразу же отметим, что процессы с участием спина — это квантовые процессы, описание которых на языке классической физики невозможно.
В биологической ткани магнитные моменты имеют разную природу. Помимо магнитных спиновых моментов электронов, это магнитные моменты протонов и других магнитных ядер и моменты орбитального движения электронов или заряженных молекулярных групп.
Энергия магнитных моментов — первичных «мишеней» МП, инициирующих биологический отклик, — чрезвычайно мала, что порождает следующее несоответствие. Энергия, например, электронного момента в геоМП равна около 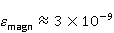 эВ. Это на семь порядков ниже высоты барьера химической активации эВ. Это на семь порядков ниже высоты барьера химической активации 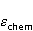 в диапазоне от единиц до десятков в диапазоне от единиц до десятков 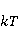 , где , где 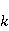 — постоянная Больцмана и — постоянная Больцмана и 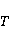 — температура среды. При физиологической температуре — температура среды. При физиологической температуре 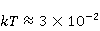 эВ. Физически, — это, по порядку величины, средняя тепловая энергия эВ. Физически, — это, по порядку величины, средняя тепловая энергия 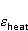 на одну степень свободы в состоянии теплового равновесия. Объединяя физический и химический аспекты, можно записать на одну степень свободы в состоянии теплового равновесия. Объединяя физический и химический аспекты, можно записать 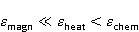 , или , или
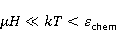
где есть температура ближайшего окружения магнитного момента (или — равновесная тепловая энергия частицы, несущей магнитный момент). Первое неравенство фиксирует несопоставимость по величине магнитных и тепловых сил, действующих на частицу. Эти неравенства породили т.н. проблему , которая долго вызывала сомнения в существовании магнитобиологических эффектов. Действительно, кажется удивительным, что МП при таком большом несоответствии может вызывать химические изменения.
Как же решается проблема ? Забегая вперед, отметим в отношении первого неравенства 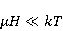 , что величина микроскопических магнитных моментов , что величина микроскопических магнитных моментов 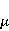 по порядку величины равна электронному магнитному моменту или на три порядка меньше у моментов ядерных. Величина МП по порядку величины равна электронному магнитному моменту или на три порядка меньше у моментов ядерных. Величина МП 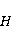 ограничена полями уровня геоМП. Поэтому их произведение ограничена полями уровня геоМП. Поэтому их произведение 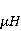 не может быть произвольно большим. Однако, — для преодоления неравенства, — может быть малой эффективная температура магнитного момента , если момент достаточно хорошо изолирован от возмущающего действия термостата. Правда, при этом второе неравенство приобретает вид не может быть произвольно большим. Однако, — для преодоления неравенства, — может быть малой эффективная температура магнитного момента , если момент достаточно хорошо изолирован от возмущающего действия термостата. Правда, при этом второе неравенство приобретает вид 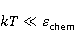 , что кажется непреодолимым препятствием для возникновения магнитных эффектов. Но это не так. Существует класс химических реакций, изучаемых спиновой химией, — реакций с участием пар радикалов [40], — в которых МП является не энергетическим, а контролирующим фактором. Механизм таких реакций рассмотрен в следующих разделах. Коротко, правила спинового отбора могут запрещать реакции, инициируемые тепловыми возмущениями среды, но запрет может быть частично снят действием МП на спиновые магнитные моменты. Поэтому именно первое неравенство является главной проблемой. Однако, как сказано выше, магнитные моменты, слабо связанные с термостатом, относительно долгое время пребывают в условиях малой эффективной температуры, — когда , что кажется непреодолимым препятствием для возникновения магнитных эффектов. Но это не так. Существует класс химических реакций, изучаемых спиновой химией, — реакций с участием пар радикалов [40], — в которых МП является не энергетическим, а контролирующим фактором. Механизм таких реакций рассмотрен в следующих разделах. Коротко, правила спинового отбора могут запрещать реакции, инициируемые тепловыми возмущениями среды, но запрет может быть частично снят действием МП на спиновые магнитные моменты. Поэтому именно первое неравенство является главной проблемой. Однако, как сказано выше, магнитные моменты, слабо связанные с термостатом, относительно долгое время пребывают в условиях малой эффективной температуры, — когда 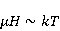 , — и магнитный эффект успевает развиться до того, как в результате релаксации установится тепловое равновесие. , — и магнитный эффект успевает развиться до того, как в результате релаксации установится тепловое равновесие.
Вышеприведенные неравенства позволяют отсеять многие гипотезы о природе магнитных биологических эффектов. Это гипотезы, которые не могут объяснить, почему слабое МП, изменяющее энергию магнитных моментов лишь на одну десятимиллионную часть , способно вызвать эффекты, преодолевающие тепловые возмущения этих магнитных моментов.
В частности, полагают иногда, что МП, воздействуя на магнитный момент микрочастицы, локализованной в каком-либо осцилляторном потенциале, могло бы привести к росту энергии осцилляций до уровня и инициировать какую-либо химическую реакцию. Такой механизм обходит проблему за счет накопления магнитной энергии в механических колебаниях. Но его реализация невозможна ввиду инерционности механических движений. Действительно, в инерционных процессах вынужденное изменение энергии механического движения или координаты частицы пропорционально квадрату времени  . Тогда обычные для магнитобиологии малые магнитные силы могут сообщить микрочастице энергию лишь за абсурдно долгое время, — даже в самой благоприятной идеализированной ситуации и в отсутствие тепловых возмущений. Поскольку механизмы магнитных эффектов, сформулированные на языке классической физики, необходимо включают механическое движение, изменяемое МП, они являются нереальными из-за инерции. Хотя в сильных МП порядка 1 Тл и более они могли бы иметь место [41], они не могут лежать в основе неспецифических эффектов слабых МП. Попытки объяснить магнитные эффекты в биологии в рамках классической физики, — иначе, чем посредством действия МП на квантовую динамику магнитных моментов, — публикуют, к сожалению, и сегодня. Важно, поэтому, подчеркнуть несостоятельность таких попыток. . Тогда обычные для магнитобиологии малые магнитные силы могут сообщить микрочастице энергию лишь за абсурдно долгое время, — даже в самой благоприятной идеализированной ситуации и в отсутствие тепловых возмущений. Поскольку механизмы магнитных эффектов, сформулированные на языке классической физики, необходимо включают механическое движение, изменяемое МП, они являются нереальными из-за инерции. Хотя в сильных МП порядка 1 Тл и более они могли бы иметь место [41], они не могут лежать в основе неспецифических эффектов слабых МП. Попытки объяснить магнитные эффекты в биологии в рамках классической физики, — иначе, чем посредством действия МП на квантовую динамику магнитных моментов, — публикуют, к сожалению, и сегодня. Важно, поэтому, подчеркнуть несостоятельность таких попыток.
Со времени появления знаменитой книги «Что такое жизнь?» [42] одного из основателей квантовой механики Э. Шредингера, обнаруживается все больше биологических процессов, изучение которых требует «квантового микроскопа». Помимо процессов, управляющих мутациями и наследственностью, это — фотосинтез, дыхание, зрение, обоняние, ферментные реакции в целом и другие формы передачи энергии и заряда. Даже для понимания такого сложного феномена, каким является сознание, сегодня привлекают идеи квантовой физики, см. обзор в [43]. По мере того, как биофизика и молекулярная биология осваивают нано и суб-нано диапазоны размеров и времен, использование квантовой физики становится неизбежным [44]. Образовалось и соответствующее понятие, — квантовая биология, — объединяющее исследования биологических процессов, описание которых на языке классической физики не продуктивно [45,46]. Магнитобиологические эффекты — тоже квантовые. Отклик организмов на слабое магнитное поле невозможно трактовать иначе, чем влияние МП на динамику магнитных моментов, главным представителем которой в организме является квантовая динамика электронов.
Следующая важная проблема состоит в том, что неспецифические эффекты трудно воспроизводимы. В отличие от специфических, неспецифические эффекты, полученные в одной лаборатории, трудно воспроизвести в другой, за что магнитобиологию часто критикуют. Снижение воспроизводимости объясняют влиянием разнообразных случайных факторов, влияющих на исход эксперимента [47].
Во-первых, значения контролируемых факторов никогда не бывают строго постоянными. Колебания МП, например, в помещениях достигают сотен нТл и даже более вблизи линий электропередач и электротранспорта [48]. Неоднородность МП в биологических термостатах доходит до десятков мкТл, что часто упускают в исследованиях магнитобиологии [49], хотя известно, что некоторые животные чувствительны к МП величиной 15–30 нТл [39,50,51] и даже 5 нТл [52]. Биологическое действие магнитных бурь подтверждено их воспроизведением в искусственных условиях [53,54] либо путем их компенсации [55]. Однако в биологических лабораториях уровень геомагнитных вариаций почти никогда не отслеживают. Отсюда случайность результатов измерений, а значит и сниженная воспроизводимость. Имеется и методологическая причина сниженной воспроизводимости — недостаточная адекватность используемых статистических методов особенностям магнитных биологических эффектов [56,57].
Во-вторых, неспецифические эффекты в магнитобиологии зависят от факторов разной природы [58–60], не все из которых поддаются контролю. Только физических факторов больше десяти [56]. Многие биологические процессы чувствительны к изменениям, например, температуры на уровне 0.1oC, хотя такие флуктуации типичны для термостатов. Кроме температуры, МП и электрического поля, это влажность, давление, освещение, скорость их изменений и др. Существуют также химические, физиологические и генотипические факторы.
Наконец, низкая воспроизводимость неспецифических эффектов имеет причиной случайность, возникающую уже на микроуровне. Это связано с тем, что преобразование сигнала МП в сигнал биохимический зависит от случайных внутренних условий.
Случайность — на фоне гомеостаза — проявляется на всех масштабах времени и уровнях организации живого вещества, начиная с молекулярного и заканчивая уровнем медленных эволюционных процессов [61] с. 281–283. Тепловые, химические и биохимические флуктуации делают случайными внутренние условия функционирования множества биофизических структур, включая биофизические сенсоры МП. Внутренние условия — это состояние локального окружения сенсоров МП. Это флуктуирующие концентрации, активности и стерические ограничения белковых агентов, особенности взаимодействий с термостатом, а также молекулярные вращения [62]. Такими условиями, например, для спин-коррелированной пары радикалов как магнитного сенсора, — см. ниже, — являются вязкость среды, детали взаимодействий, определяющих время тепловой релаксации спинов, скорость химической реакции радикалов, концентрации и др.
Внутренние условия в организме непрерывно меняются вследствие регуляторных процессов, контролируемых множеством рецепторных систем и обратных связей. Измерения биологических характеристик попадают в разные фазы регуляторных процессов. Это означает, что отклик на изменение МП случаен из-за флуктуаций внутренних условий, вызванных как внешними, так и внутренними регуляторными факторами. Флуктуации условий постоянно «включают» и «выключают» различные группы магнитных биофизических сенсоров. Следовательно, результаты измерений, полученные с разными группами включенных сенсоров, формируют даже не просто случайную величину, а совокупность реализаций разных случайных величин, т.е. гетерогенный случайный эффект. Добавим, что в отличие от специализированных рецепторов, локализованных в определенных участках организма, магнитные моменты электронов и ядер, спиновые и орбитальные, присутствуют практически во всех биологических молекулах, распределенных по всему организму. Можно ли заранее сказать, какие из них передадут сигнал МП на уровень биохимических реакций и где это произойдет? Пока нет. В этих условиях — при гетерогенности откликов — понятия порога, дозы, да и МП-зависимости вообще, имеют лишь ограниченный смысл, что говорит о необходимости специальных методов изучения неспецифических эффектов [56].
Наконец, отметим еще одну проблему магнитобиологии неспецифических эффектов — проблему биологического детектирования малых вариаций МП на фоне действия гораздо бóльших случайных флуктуаций магнитного поля в городских условиях. Без решения этой проблемы невозможно объяснить, например, действие геомагнитных бурь, интенсивностью редко превышающих 500 нТл, на функционирование организмов. Биологические эффекты магнитных бурь зафиксированы как в организме человека, так и в других организмах [63,64]. Низкочастотные флуктуации геоМП, как и городского магнитного шума, оказывают на организмы, по-видимому, непосредственное действие [30,51,53,65].
Скорее всего, эти факты не потребуют привлечения каких-либо дополнительных объяснительных механизмов. Объяснение возможно на единой основе влияния МП на магнитные моменты. Следует лишь учесть частотный и временной факторы магнитной экспозиции. Так, с одной стороны, в [36] отмечено, что частотные спектры гео- и городских магнитных флуктуаций смещены относительно друг друга на два-три порядка. Предположено, что процесс формирования биологического отклика на первичные сигналы геомагнитных возмущений в организме включает их интегрирование в системе биохимических сигнализаторов и регуляторов. Концентрация молекул, биосинтез и распад которых медленен, может отражать накопленное и усредненное действие слабых магнитных сигналов. С другой стороны, в [48] показано, что интенсивность геомагнитного шума в низкочастотной области может превышать интенсивность городского магнитного шума в ночные часы, когда и наблюдаются, главным образом, неблагоприятное влияние магнитных бурь на состояние человека.
Какие механизмы наиболее правдоподобны?
Малая эффективная температура, или метастабильность, первичной мишени и безынерционность ее отклика на МП как условие работы молекулярных механизмов действия слабых МП на биологические системы являются существенными ограничителями. Правдоподобные механизмы должны опираться, во-первых, на динамику магнитных моментов. Действительно, конечная угловая скорость прецессии магнитных моментов пропорциональна приложенному МП и не зависит от времени явно. Прецессия момента возникает в МП немедленно и не требует накопления энергии микрочастицы. Это следствие независимости энергии спина или механического вращения, связанного с магнитным моментом, от направления момента. Направление углового момента меняется здесь пропорционально , т.е. безынерционно [47]. Во-вторых, динамика магнитных моментов должна быть квантовой, поскольку освободиться от вовлеченности спинов в магнитные эффекты невозможно. С другой стороны, если говорить об угловых моментах орбитального движения, то они очень малы, того же порядка, что и спин электрона, и их квантовый характер следует из соотношения неопределенности Гейзенберга. В-третьих, прецессия моментов должна происходить в условиях медленной тепловой релаксации, когда действием тепловых возмущений можно пренебречь. Поэтому к правдоподобным относятся механизмы влияния МП на абстрактные одиночные магнитные моменты [66], на спиновые магнитные моменты электронов радикальных пар [28,32,67], на магнитные моменты ядер [68,69] и орбитального движения молекулярных групп внутри белков и в других биофизических структурах [27,70]. Медленность тепловой релаксации, или относительно большой временной интервал когерентности движения моментов, в этих механизмах пока не доказана, а лишь предполагается. Подробнее об этом будет сказано ниже.
Постепенно развивается предположение о том, что протонный обмен и метастабильные состояния в жидкой воде могли бы отвечать за неспецифические магнитные эффекты [71–73]. Однако эти гипотетические механизмы еще не приобрели форму математических моделей, обладающих предсказательной силой. Пока они не удовлетворяют одному из принципов научной методологии — принципу проверяемости. Поэтому мы ограничимся обсуждением достоинств и недостатков вышеуказанных механизмов действия МП на магнитные моменты в биофизических структурах. Эти механизмы возникают естественным образом при рассмотрении общих свойств химического процесса.
Неспецифическое действие слабых МП — это, по определению, влияние МП на химический процесс в отсутствие рецепторов, т.е. в достаточно общих условиях. В химическом процессе выделяют две основные стадии: 1) сближение реагентов, часто диффузионное, на расстояние порядка длин химических связей и 2) собственно акт химической реакции, т.е. перегруппировка электронных оболочек. При физиологической температуре энергия, сообщаемая магнитными силами, на много порядков меньше энергии диффузионного движения. По этой причине МП не может влиять на ход процесса на первой стадии.
Однако МП способно влиять на квантовый процесс второй стадии — на акт реакции. Априори влияние могло бы осуществляться по-разному. Влияние могло бы происходить посредством спинового эффекта на вероятность перегруппировки электронов. Другой путь осуществляется через один из стерических факторов, через влияние МП на пространственную структуру электронных волновых функций взаимодействующих молекул. Таким образом, если ключевым фактором является перестройка электронных оболочек под действием МП, то правдоподобны следующие сценарии.
 МП влияет на перегруппировку электронов, например, в реакции распада молекулы МП влияет на перегруппировку электронов, например, в реакции распада молекулы 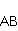 на свободные радикалы на свободные радикалы 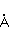 и и 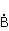 , если реакция идет с образованием интермедиата , если реакция идет с образованием интермедиата 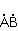
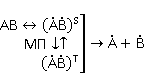
который может находиться в разных спиновых состояниях. Это простой пример реакций с участием радикальных пар. Такие реакции изучает спиновая химия.
МП влияет на прецессию (в векторной модели) одиночных моментов, например, спиновых магнитных моментов электронов. Прецессия становится неравномерной в переменном МП или замедляется в гипомагнитном МП [66], что может сказаться на вероятности актов смежных реакций, т.е. также повлиять на перегруппировку электронов.
МП изменяет пространственную структуру или фазы угловых квантовых состояний вращающихся молекулярных групп и электронных облаков, что приводит к интерференционным эффектам [74].
В отличие от первого сценария, или спин-химического механизма радикальных пар (РПМ), — последние два — механизм смешения квантовых уровней [66] и механизм молекулярного гироскопа [74] — на сегодня не имеют надежного экспериментального подтверждения. Хотя они и предсказывают ряд наблюдаемых особенностей неспецифических эффектов, которые РПМ объяснить не может, они, тем не менее, остаются умозрительными конструкциями. Начальная информация о них дана в Приложениях 1 и 2, а ниже мы продолжаем рассматривать РПМ, — наиболее правдоподобный сценарий неспецифических магнитных эффектов. Радикальные пары возникают, например, в фотолиазах и криптохромах под действием оптического синего и УФ излучения. Возникают они и во многих фермент-субстратных [75,76], например, биополимерных [40], реакциях, поскольку в активном центре фермента часто имеет место окислительно-восстановительный процесс, сопровождающийся переносом электрона. Все это создает перспективную основу для объяснения магнитных эффектов в организмах.
Поясним процессы, изображенные на вышеприведенной схеме возникновения свободных радикалов. Пусть часть 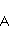 сложной молекулы содержит ковалентную связь, разрываемую в результате переноса электрона на часть сложной молекулы содержит ковалентную связь, разрываемую в результате переноса электрона на часть 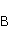 . В самом простом случае разрывается ковалентная связь, соединяющая и . Сразу после переноса электрона образуются два радикала, и , поскольку части молекулы содержат теперь неспаренные электроны. Спиновое состояние радикальной пары в начальный момент времени является коррелированным: спины электронов, локализованных на и , эволюционируют согласованно, или когерентно, так как случайные тепловые возмущения еще не успели разрушить когерентность. . В самом простом случае разрывается ковалентная связь, соединяющая и . Сразу после переноса электрона образуются два радикала, и , поскольку части молекулы содержат теперь неспаренные электроны. Спиновое состояние радикальной пары в начальный момент времени является коррелированным: спины электронов, локализованных на и , эволюционируют согласованно, или когерентно, так как случайные тепловые возмущения еще не успели разрушить когерентность.
Для описания спиновых состояний обычно используют т.н. зеемановский базис, 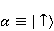 и и 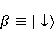 , обозначающий состояния с проекцией спина на ось МП , обозначающий состояния с проекцией спина на ось МП 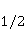 и и 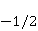 , соответственно. Одно из базисных состояний системы двух электронов обозначается тогда как , соответственно. Одно из базисных состояний системы двух электронов обозначается тогда как 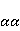 , где на первом месте указано спиновое состояние первого электрона, а на втором месте — второго. Другие базисные состояния пары есть, очевидно, , где на первом месте указано спиновое состояние первого электрона, а на втором месте — второго. Другие базисные состояния пары есть, очевидно, 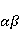 , , 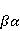 и и 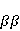 . .
Стационарные состояния пары — это собственные состояния оператора энергии — гамильтониана системы. Стационарные состояния обладают определенной симметрией по перестановке частиц. Если имеется только энергия магнитных моментов в МП, то это антисимметричное синглетное состояние 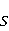 , в котором суммарный спин пары равен нулю, , в котором суммарный спин пары равен нулю, 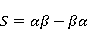 , и три триплетных состояния , соответствующие суммарному спину единица с разными проекциями спина на направление МП. Триплетные состояния симметричны по перестановке пары частиц: , и три триплетных состояния , соответствующие суммарному спину единица с разными проекциями спина на направление МП. Триплетные состояния симметричны по перестановке пары частиц: 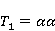 , , 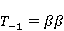 и и 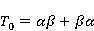 . .
Перенос электрона с одной части молекулы на другую, — или с одной молекулярной группы на другую, как в белках, — означает появление двух парамагнитных электронов в синглетном совместном состоянии. Это есть следствие закона сохранения углового момента системы двух электронов (напомним, что спин — это собственный угловой механический момент частицы). Суммарный угловой момент пары электронов в результате переноса как правило не меняется, а до переноса он был нулевым, так как принадлежал спаренным электронам ковалентной связи.
В некоторых случаях интермедиат , возникающий в результате переноса электрона и первоначально находящийся в синглетном состоянии 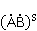 , живет достаточно долго, и МП может повлиять на эволюцию его спинового состояния, или, что то же, изменить скорость синглет-триплетных (СТ) переходов , живет достаточно долго, и МП может повлиять на эволюцию его спинового состояния, или, что то же, изменить скорость синглет-триплетных (СТ) переходов 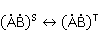 , происходящих по разным причинам. Поскольку рекомбинация частей интермедиата в начальную форму возможна только из синглетного состояния, СТ переходы, сперва уменьшающие долю синглетного состояния, уменьшают вероятность рекомбинации и, значит, увеличивают вероятность образования свободных радикалов и . Это послужило основанием для частых предположений, что МП увеличивает концентрацию свободных радикалов в организме и ведет к разнообразным негативным эффектам. Однако процессы переноса с возникновением радикальных пар в организме сложнее, чем вышеприведенная иллюстративная схема, а возможный относительный рост концентрации свободных радикалов, если и происходит, то очень мал и не объясняет наблюдаемого влияния слабых МП на организмы. Это будет пояснено ниже. , происходящих по разным причинам. Поскольку рекомбинация частей интермедиата в начальную форму возможна только из синглетного состояния, СТ переходы, сперва уменьшающие долю синглетного состояния, уменьшают вероятность рекомбинации и, значит, увеличивают вероятность образования свободных радикалов и . Это послужило основанием для частых предположений, что МП увеличивает концентрацию свободных радикалов в организме и ведет к разнообразным негативным эффектам. Однако процессы переноса с возникновением радикальных пар в организме сложнее, чем вышеприведенная иллюстративная схема, а возможный относительный рост концентрации свободных радикалов, если и происходит, то очень мал и не объясняет наблюдаемого влияния слабых МП на организмы. Это будет пояснено ниже.
В оценке правдоподобности РПМ важным является учет тепловых возмущений, которые разрушают согласованность, или когерентность, динамики магнитных моментов или спинов электронов. СТ состояния электронных спинов имеют смысл (описание в терминах СТ состояний эффективно) только если динамика спинов когерентна. Когерентность разрушается тепловыми флуктуациями. В термализованном состоянии движение спинов существенно хаотично. МП не способно повлиять на эволюцию пары спинов, или на СТ конверсию. Магнитные эффекты возможны только на интервале когерентности спинов, или в течение времени  релаксации спиновой когерентности, рис. 4. релаксации спиновой когерентности, рис. 4.
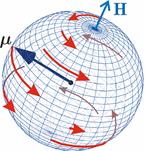
Рис. 4. Характер прецессии «вектора» спинового магнитного момента 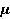 в МП в МП 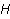 под действием тепловых возмущений. Красные стрелки — траектории движения конца вектора в промежутках между тепловыми ударами. На этих участках прецессия является когерентной. В конце каждого такого участка происходит быстрый случайный перескок вектора в начало какого-либо другого участка. Если тепловые возмущения малы, то удары редки, и прецессия главным образом когерентна. Если возмущения велики, то участки когерентности очень малы и движение неотличимо от случайного. В таком состоянии МП не влияет на эволюцию магнитного момента. Следует иметь в виду, что векторная модель спина является лишь удобной иллюстрацией квантовомеханического процесса, поскольку вектора спина как такового не существует. под действием тепловых возмущений. Красные стрелки — траектории движения конца вектора в промежутках между тепловыми ударами. На этих участках прецессия является когерентной. В конце каждого такого участка происходит быстрый случайный перескок вектора в начало какого-либо другого участка. Если тепловые возмущения малы, то удары редки, и прецессия главным образом когерентна. Если возмущения велики, то участки когерентности очень малы и движение неотличимо от случайного. В таком состоянии МП не влияет на эволюцию магнитного момента. Следует иметь в виду, что векторная модель спина является лишь удобной иллюстрацией квантовомеханического процесса, поскольку вектора спина как такового не существует.
В теории спиновой релаксации различают времена продольной и поперечной релаксации. Нас интересуют порядки величин, поэтому мы используем обобщающее понятие характерного времени тепловой спиновой релаксации . В большинстве РМП моделей магнитной навигации животных время спиновой релаксации полагается очень большим, — больше, чем все другие характерные временные масштабы релевантных процессов. Поэтому эффекты термализации попросту не учитывают. Поскольку основания к такой идеализации весьма спорны, мы далее обсуждаем вклад термализации спинов подробнее. Но сначала сделаем одно существенное уточнение в отношении описания квантовых систем в терминах СТ состояний.
В эксперименте наблюдали повышение концентрации активных форм кислорода, в частности синглетного кислорода, при действии относительно слабых МП. В отличие от триплетного основного состояния молекулярного кислорода, его синглетная форма с наименьшей энергией, называемая синглетным кислородом, обладает высокой реакционной способностью и может повреждать биополимеры [77]. По этой причине предполагают иногда, что МП влияет на организм, непосредственно возбуждая СТ переходы и повышая концентрацию синглетного кислорода. Это предположение неверно. Разность энергий С- и T-состояний молекулы кислорода соответствует энергии квантов излучения ближнего ИК-диапазона около 1280 нм, что слишком много для возникновения эффектов слабого МП.
Рассмотрим это обстоятельство подробнее. СТ состояния молекулярных термов существенно отличаются от СТ состояний пары радикалов, что есть следствие огромной разницы в величине обменного взаимодействия электронов в молекуле кислорода и в типичной радикальной паре в белках. Электроны радикалов в белках разнесены в пространстве на расстояние порядка 1 нм, и короткодействующее обменное взаимодействие мало, поскольку его магнитуда определяется перекрытием волновых функций двух электронов. Волновые функции экспоненциально спадают с ростом расстояния между электронами, и для парамагнитных электронов в белках обменным взаимодействием обычно пренебрегают. В молекуле кислорода два электрона локализованы около одного и того же ядра, поэтому их волновые функции существенно перекрываются. Здесь обменное взаимодействие электронов велико и приводит — согласно принципу запрета Паули — к значительному энергетическому зазору между С- и Т-термами, порядка 1 эВ. Вследствие этого магнитные эффекты в отношении СТ состояний молекулярного кислорода начинают проявляться в МП порядка 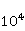 Э ( Э (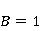 Тл) [78], когда энергия магнитного момента электрона сравнима с величиной зазора. Слабые МП порядка геоМП на 4–5 порядков меньше и прямого влияния на молекулу кислорода не оказывают. Тл) [78], когда энергия магнитного момента электрона сравнима с величиной зазора. Слабые МП порядка геоМП на 4–5 порядков меньше и прямого влияния на молекулу кислорода не оказывают.
Механизм радикальных пар
В отличие от влияния сильного МП на молекулу кислорода, влияние МП на пару радикалов промежуточного состояния имеет место в гораздо меньших полях, начиная с нескольких мТл. Поэтому спиново-химический механизм, РПМ, считают сегодня основным кандидатом на объяснение эффектов слабых МП.
Его главное достоинство в том, что он безусловно существует и удачно согласуется с некоторыми наблюдаемыми в магнитной навигации закономерностями. Это следующие закономерности: 1) зависимость отклика на МП от присутствия оптического излучения и его спектрального состава, 2) инвариантность к изменению МП на противоположно направленное и 3) отсутствие выраженной частотной селективности в НЧ диапазоне переменного МП. Полагают, что излучение из синей области оптического спектра приводит к фотоиндуцированному забросу электрона на один из верхних уровней, за которым следует его перенос на другой центр и, таким образом, рождение электронной пары радикалов. Квантово-механическая модель РПМ предсказывает, что эффект является функцией квадрата величины МП [79] и, значит, инвариантен к инверсии МП. Главный недостаток РПМ — его низкая чувствительность к МП. Это пока мешает безоговорочно признать, что спин-химический механизм объясняет отклик организмов на МП уровня геоМП и, тем более, уровня десятков нТл.
Выше отмечалось, что магнитное чувство некоторых животных совмещено со зрительным аппаратом. Рассмотрим подробнее, каким образом РПМ встроен в зрительную систему. Зрительный фоторецептор есть расположенная в сетчатке глаза и знакомая со школы «колбочка» или «палочка», содержащая в себе белок криптохром. С одной стороны, при помощи этого белка происходит преобразование светового сигнала в биохимический. С другой стороны, такое преобразование включает рождение пар радикалов, а они чувствительны к МП. За миллиарды лет эволюции фоторецепторы некоторых птиц стали участвовать в магниторецепции [32]. Полагают, что фотоны синего спектрального диапазона инициируют рождение флавин-триптофановых радикальных пар, спиновая, а следовательно, и химическая эволюция которых зависит от МП. Многие подробности еще не известны, но сам факт подтвержден многочисленными публикациями в высокорейтинговых журналах, см. напр. [33,80,81].
Объяснение действия МП на фотосинтетические бактерии предложено в [82]. Населенность триплетного состояния пары фотоиндуцированных радикалов в реакционном центре зависит от величины МП. Это и влияет на фотосинтез. Годом позже авторы [79] предположили, что ориентирование птиц в геоМП происходит при помощи квантового химического компаса. СТ конверсия радикалов с учетом анизотропного сверхтонкого взаимодействия зависит также и от направления (но не от реверса) МП, что согласуется с экспериментом [21]. Подобные механизмы получили коллективное название РПМ. Данный механизм, однако, обладает низкой чувствительностью. В отдельном фоторецепторе изменение МП на величину порядка геоМП вызывает эффект, вряд ли превышающий 0.1% теоретически, а экспериментально — на порядки меньше [62,83].
Низкая чувствительность РПМ обусловлена малым временем жизни коррелированного состояния спинов — 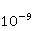 с, редко с, редко 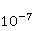 с. Это время тепловой релаксации фазовой когерентности электронных спинов. За малое время МП должно успеть заметно изменить состояние пары спинов относительно друг друга. Оценить магнитуду этого МП можно, сравнивая ширину квантового уровня, связанную со временем тепловой релаксации, и размер зеемановского расщепления уровней в МП. В [27,66] показано, что качественные изменения в динамике моментов происходят когда зазор с. Это время тепловой релаксации фазовой когерентности электронных спинов. За малое время МП должно успеть заметно изменить состояние пары спинов относительно друг друга. Оценить магнитуду этого МП можно, сравнивая ширину квантового уровня, связанную со временем тепловой релаксации, и размер зеемановского расщепления уровней в МП. В [27,66] показано, что качественные изменения в динамике моментов происходят когда зазор 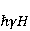 между зеемановскими подуровнями становится сравним с шириной между зеемановскими подуровнями становится сравним с шириной 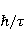 самих уровней. Следовательно, чувствительность биофизического сенсора на основе РПМ может возникать там, где имеет место фундаментальное соотношение самих уровней. Следовательно, чувствительность биофизического сенсора на основе РПМ может возникать там, где имеет место фундаментальное соотношение 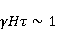 . Параметр . Параметр 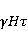 , в числе многих других, управляет вероятностью реакции сенсора на МП. Таким образом МП, начиная с которого могут стать заметны магнитные эффекты, — это , в числе многих других, управляет вероятностью реакции сенсора на МП. Таким образом МП, начиная с которого могут стать заметны магнитные эффекты, — это 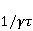 , где — время тепловой релаксации момента. Для электронов с , где — время тепловой релаксации момента. Для электронов с 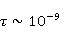 с, — например, у радикалов фермент-субстратных комплексов, — эта величина составляет 5 мТл, что в сто раз больше, чем геоМП. Для чувствительности хотя бы на уровне 5 мкТл, наблюдаемой в неспецифических эффектах, время релаксации фазовой когерентности спинов радикальной пары должно превышать 1 мкс! Пока неясно, могли бы вообще в живой ткани существовать условия, обеспечивающие столь слабую связь электрона с термостатом [84,85]. Некоторые авторы полагают [86], что наблюдение биологической реакции в столь слабых МП доказывает, что длительные времена спиновой релаксации электронов действительно существуют. с, — например, у радикалов фермент-субстратных комплексов, — эта величина составляет 5 мТл, что в сто раз больше, чем геоМП. Для чувствительности хотя бы на уровне 5 мкТл, наблюдаемой в неспецифических эффектах, время релаксации фазовой когерентности спинов радикальной пары должно превышать 1 мкс! Пока неясно, могли бы вообще в живой ткани существовать условия, обеспечивающие столь слабую связь электрона с термостатом [84,85]. Некоторые авторы полагают [86], что наблюдение биологической реакции в столь слабых МП доказывает, что длительные времена спиновой релаксации электронов действительно существуют.
Авторы [87,88] пытались обосновать большие времена спиновой релаксации теоретически. Однако до сих пор нет экспериментов, подтверждающих существование такого большого времени релаксации, за исключением экзотических систем типа фуллеренов, не имеющих отношения к биологии. Действительное время спиновой релаксации в радикальных парах может быть получено из измерений ширины линии ЭПР в геоМП или измерениями спиновых магнитных эффектов in vitro и их сравнением с расчетами. Насколько нам известно, сигналы ЭПР в геоМП от электронных спинов промежуточных радикальных пар в биохимических реакциях не наблюдались. В то же время опыт спиновой химии, см. напр. [83], согласующийся с теорией, указывает на относительно быструю спиновую релаксацию в обсуждаемых условиях, около 1 нс. Возможно, самое большее, 10 нс. Магнитные эффекты уменьшаются примерно пропорционально квадрату времени спиновой релаксации, — если скорость химической реакции не велика и не подавляет магнитный эффект в еще большей степени. Поэтому вопрос о времени релаксации спиновой когерентности эквивалентен вопросу о существовании магнитных эффектов по механизму РПМ в его стандартной форме.
Спиновая когерентность является одним из обязательных условий работы квантовых компьютеров, — условием, ограничивающим их эффективность. Поэтому некоторые авторы изучают уместность описания РПМ-магниторецепции в терминах, типичных для квантовых компьютеров, — запутанности квантовых состояний и различения между чистыми и смешанными состояниями [89]. Пока неясно, может ли такое расширение терминологической базы дать что-то новое для решения проблем магнитобиологии.
Концепция магнитной навигации на основе спин-химических эффектов в криптохромах предсказывает эффекты качественно согласующиеся со значительной частью наблюдений и экспериментов. На данный момент эта концепция является вполне обоснованной гипотезой, не лишенной, однако, вышеуказанной трудности.
Статистическое усиление слабых магнитных сигналов
Как следует из представленного выше материала, спиновая химия, — или, в более общем контексте, динамика магнитных моментов, — с необходимостью лежит в основе наблюдаемых биологических эффектов слабых МП. С другой стороны, анализ вышеприведенных квантовых механизмов, показывает, что величина первичных физических изменений, в частности в РПМ, недостаточна для надежного объяснения результатов экспериментов и наблюдений.
По-видимому, малые первичные изменения, происходящие вследствие действия МП, должны быть каким-то образом «усилены», чтобы затем вызвать ощутимые изменения в организме на уровне концентраций биохимических агентов. Данная идея не нова. В качестве усилительного механизма предлагали ферментативную реакцию, высокочувствительную к концентрации фермента [90] и химическую реакцию в режиме, близком к бифуркационной неустойчивости, когда путь реакции может измениться скачком при незначительной вариации концентрации реактанта [91–93]. Эти подходы не получили развития, — вероятно вследствие невозможности их экспериментальной верификации in vivo. Для объяснения магнитного чувства некоторых животных привлекали идею статистического усиления одновременно поступающих в мозг очень малых магнитных сигналов от миллионов фоторецепторов. Коэффициент усиления оказался недостаточным для объяснения магнитной навигации животных на основе РПМ [94]. Тем более этого механизма не достаточно в отношении неспецифических эффектов, — в отсутствие эволюционно развитых магниторецепторов.
Однако, как отмечено выше, сама концепция статистического усиления, или накопления, малых регулярных отклонений на фоне флуктуаций привлекательна. Эта концепция детектирования слабых сигналов работает везде, где уместны понятия случайной величины, ее среднего и вариаций, — она общеизвестна и универсальна. Следовало бы ожидать, что магнитные эффекты, если они обусловлены первичными малыми отклонениями под действием МП, могли бы быть обнаружены там, где они накапливаются в длинной последовательности элементарных актов.
Обратим внимание на то, что циклические процессы синтеза биополимеров являются идеальными биохимическими интеграторами, так что малые изменения в отдельных циклах реакций могут накапливаться и приводить к наблюдаемым биологическим эффектам. Биологический отклик на первичные магнитные сигналы возникает благодаря интегрированию в биохимической системе организма. Особый интерес связан с возникновением ошибок в этих процессах. Даже одна ошибка в каком-либо отдельном цикле может привести к неработоспособности целой молекулы синтезированного биополимера. Покажем, как малый первичный магнитный сигнал мог бы быть статистически усилен вместе с накоплением вероятности ошибок биополимеризации [95].
Процессы синтеза биополимеров разнообразны. Это репликация ДНК, транскрипция — синтез комплементарной РНК, сплайсинг, трансляция — синтез белка из аминокислот в соответствии с кодом мРНК и посттрансляционный фолдинг белковой цепи в глобулу и ее созревание. На каждом из этих этапов могут происходить случайные ошибки, однако с существенно разной вероятностью. Имеются совершенные биохимические механизмы коррекции ошибок репликации и транскрипции; их вероятность поэтому очень мала, порядка 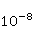 и и 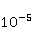 , соответственно [96]. Ошибки трансляции случаются гораздо чаще, с вероятностью порядка , соответственно [96]. Ошибки трансляции случаются гораздо чаще, с вероятностью порядка 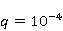 – –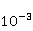 на одну присоединенную молекулу транслируемой аминокислоты [97,98]. Около десяти процентов синтезируемых молекул из нескольких сотен звеньев содержат хотя бы одну ошибку. Для таких молекул шанс принять в процессе фолдинга нативную конформацию существенно снижен. на одну присоединенную молекулу транслируемой аминокислоты [97,98]. Около десяти процентов синтезируемых молекул из нескольких сотен звеньев содержат хотя бы одну ошибку. Для таких молекул шанс принять в процессе фолдинга нативную конформацию существенно снижен.
Ошибки возникают и на стадии фолдинга, — одной из причин является запутанная геометрия и топологические узлы траекторий фолдинга [99]. Полагают, однако, что более вероятен мисфолдинг, происходящий из-за ошибок трансляций, а не собственно фолдинга. Следовательно, главным образом ошибки трансляции генетической информации контролируют точность экспрессии генов. Очевидно, что возможное влияние МП на вероятность ошибок трансляции — это именно тот процесс, где магнитные эффекты могли бы проявиться на биологическом уровне. Об этом же косвенно свидетельствуют и результаты работы [100], где слабое электромагнитное излучение влияло на экспрессию генов, не вызывая при этом разрывов ДНК.
Недавно предложен такой механизм статистического усиления, основанный на том, что МП могло бы влиять не на концентрацию нативных белков, необходимых для функционирования организма, а на концентрацию нефункциональных белков, возникающих в результате ошибок синтеза биополимеров [95]. Нефункциональные белки бывают цитотоксичными [101] и в любом случае вызывают дополнительную нагрузку на адаптивные ресурсы организма.
Трансляция — сложный многостадийный циклический процесс, включающий разнообразные ферментные реакции. Трансляция осуществляется рибосомой — специальной макромолекулярной машиной для сборки аминокислот в белки. Упрощение, иллюстрирующее возникновение заметной ошибки, состоит в следующем. Пусть имеется процесс синтеза белковой цепи из большого числа  звеньев с равной вероятностью звеньев с равной вероятностью 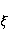 ошибки включения — появления в каком-либо месте синтезируемой цепи неправильного аминокислотного остатка, — не соответствующего инструкции мРНК. Нативная функциональная белковая глобула подразумевает отсутствие локальных ошибок во всех местах цепи. Тогда вероятность ошибки включения — появления в каком-либо месте синтезируемой цепи неправильного аминокислотного остатка, — не соответствующего инструкции мРНК. Нативная функциональная белковая глобула подразумевает отсутствие локальных ошибок во всех местах цепи. Тогда вероятность 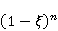 есть вероятность появления «правильной» аминокислотной последовательности, а есть вероятность появления «правильной» аминокислотной последовательности, а 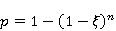 есть вероятность появления дефектной молекулы, т.е. вероятность ошибки трансляции. Чувствительность величины есть вероятность появления дефектной молекулы, т.е. вероятность ошибки трансляции. Чувствительность величины  к вероятности локальной ошибки включения — это производная к вероятности локальной ошибки включения — это производная 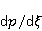 . Ее магнитуда максимальна при условии . Ее магнитуда максимальна при условии 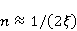 и может достигать больших значений и может достигать больших значений 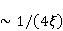 при малых . Вероятность правильной трансляции всей молекулы в любом случае не превышает единицы. Это означает, что эффект изменения вероятности ошибки трансляции при варьировании вероятности локального сбоя состоит в том, что изменение на несколько десятых долей возникает уже при вариации очень малых при малых . Вероятность правильной трансляции всей молекулы в любом случае не превышает единицы. Это означает, что эффект изменения вероятности ошибки трансляции при варьировании вероятности локального сбоя состоит в том, что изменение на несколько десятых долей возникает уже при вариации очень малых 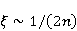 . .
Число различных белков в организме человека превышает два миллиона [102], а основная их масса обладает длиной от ста до пятисот аминокислот. Поэтому изменения вероятности локальных сбоев 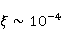 – вызывают существенные изменения вероятности появления нефункциональных белков. Заметим, что уровни – вызывают существенные изменения вероятности появления нефункциональных белков. Заметим, что уровни 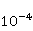 – — это обычная величина относительных эффектов в РПМ. С одной стороны, рибосомальная трансляция осуществляется с помощью специальных ферментов, а многие ферменты работают с выраженной стадией переноса электрона. С другой стороны, при переносе электрона может возникать промежуточное магниточувствительное состояние пары радикалов со спин-коррелированными электронами. Действительно, активность некоторых ферментов зависит от МП, см. напр. [76,103]. Таким образом предположение о вовлеченности РПМ или, в общем случае, динамики магнитных моментов, в рибосомальную трансляцию хорошо обосновано, а механизм статистического усиления ошибок трансляции непротиворечиво объясняет неспецифическое влияние слабых МП на организм, рис. 5. – — это обычная величина относительных эффектов в РПМ. С одной стороны, рибосомальная трансляция осуществляется с помощью специальных ферментов, а многие ферменты работают с выраженной стадией переноса электрона. С другой стороны, при переносе электрона может возникать промежуточное магниточувствительное состояние пары радикалов со спин-коррелированными электронами. Действительно, активность некоторых ферментов зависит от МП, см. напр. [76,103]. Таким образом предположение о вовлеченности РПМ или, в общем случае, динамики магнитных моментов, в рибосомальную трансляцию хорошо обосновано, а механизм статистического усиления ошибок трансляции непротиворечиво объясняет неспецифическое влияние слабых МП на организм, рис. 5.
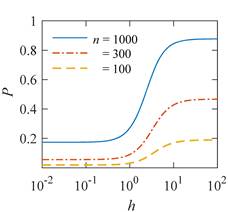
Рис. 5. Пример расчета зависимости средней вероятности 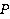 ошибки рибосомальной трансляции от относительной величины МП ошибки рибосомальной трансляции от относительной величины МП 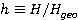 при разных размерах при разных размерах 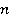 синтезируемой белковой цепи, по данным [95]. синтезируемой белковой цепи, по данным [95].
Данный механизм статистического усиления снимает основное противоречие в объяснении неспецифических магнитных эффектов на основе РПМ — очень малую величину РМП эффектов порядка –. Столь незначительные первичные магнитные эффекты получают усиление приблизительно в 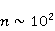 – –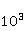 раз и выводят суммарный эффект на наблюдаемый уровень. раз и выводят суммарный эффект на наблюдаемый уровень.
Гипомагнитное поле как инструмент исследования
Исследование биополимерных процессов в гипомагнитном поле (гипоМП — МП, сниженное по сравнению с геоМП на два порядка и более) является перспективным инструментом для выявления молекулярного механизма влияния низкоинтенсивных МП на организмы и последующего использования прецизионных слабых МП для селективного управления экспрессией генов. Действительно, основа такого влияния — это первичное действие МП на магнитные моменты атомно-молекулярного масштаба. С одной стороны, на сегодня первичная мишень МП в организмах или водных системах не установлена. Рис. 5, например, демонстрирует ожидаемые зависимости вероятности эффекта по механизму РПМ, т.е. когда первичные мишени — это электроны. С другой стороны, мишенями могли бы быть и магнитные моменты протонов или других магнитных ядер, как и магнитные моменты молекулярных вращений заряженных групп. Эти типы моментов обладают разным гиромагнитным отношением и, — в случае их прецессии в МП, — прецессируют с разной угловой скоростью. Поэтому наилучшим инструментом магнитного влияния является то, которое затрагивает все магнитные моменты независимо от их гиромагнитного фактора. Это и есть гипоМП. В таком поле замедляется прецессия всех магнитных моментов. Квантовые механизмы эффектов гипоМП с участием магнитных моментов с разным гиромагнитным отношением рассмотрены в [27] и демонстрируют богатые особенностями зависимости от МП.
Однако проведение экспериментов с использованием гипоМП в обычных лабораторных условиях затруднено, поскольку флуктуации МП в лабораториях нередко достигают нескольких мкТл. Здесь имеется аналогия с применением протонного ЯМР в жидкости, когда ширина линии резонанса очень узкая, составляет – Гс. В геоМП с флуктуациями случайное смещение линии существенно превышало бы ширину линии, что затрудняло бы наблюдение резонанса. Эта трудность снимается увеличением МП на 6 порядков, то есть использованием в ЯМР полей порядка 1–10 Т, когда флуктуации МП в несколько мкТл практически не смещают линий — не вызывают заметных относительных изменений.
Другой очевидный способ снижения влияния флуктуаций МП — это активная их компенсация или экранирование, рис. 6. Именно на фоне гипоМП было бы возможно наблюдение действия дополнительно организованных прецизионных МП на молекулярные процессы в организмах. Наблюдение таких эффектов в сильных полях невозможно, так как в них прецессия всех магнитных моментов высокочастотная и не соответствует временному масштабу естественных биополимерных процессов. ГипоМП могло бы на практике быть столь же интересным, сколь и сильное.

Рис. 6. Один из двух одинаковых блоков системы гипомагнитной экспозиции, реализованной в ИОФ РАН, компенсирует внешнее МП и позволяет снизить флуктуации однородного окружающего МП в несколько тысяч раз.
Инструментальный потенциал гипоМП в сравнении с сильным МП можно представить по аналогии с использованием сильного и слабого оптического излучения. Естественным «биологическим» репером является интенсивность дневного света. Мощный свет в форме, например, лазерного излучения находит множество очевидных применений. Очень слабые излучения — одиночные фотоны — позволяют исследовать фундаментальные квантовые процессы и находят применение в разработке квантовых компьютеров и квантовой криптографии. ГипоМП — это, по-видимому, важная область биофизических исследований. Применение гипоМП полей в молекулярной биологии открывает интересные перспективы, сравнимые по универсальности с ЯМР. Если представленный выше механизм статистического усиления индуцированных МП ошибок трансляции пройдет экспериментальную апробацию и окажется верным, то станет реальным и селективное управление скоростью производства белков без использования химических препаратов — с помощью специально организованного прецизионного МП на фоне гипоМП. Это имело бы ясные перспективы применения, помимо исследовательского, в медицине, фармакологии и сельскохозяйственных биотехнологиях.
Заключение
Квантовые неспецифические эффекты слабых МП обладают общим и универсальным характером — имеют место всегда и во всех организмах. Эти эффекты незаметны, когда малы. Когда они не малы и могли бы быть заметны, они, в силу своей случайности, маскируются под результат действия неконтролируемых случайных факторов. Это проявляется в виде невоспроизводимости, — гетерогенности массива измерений или специфически увеличенного случайного разброса. В отличие от магниторецепции — действия слабых МП на специализированные рецепторы некоторых организмов, — магнитные неспецифические эффекты недополучают внимание исследователей. Молекулярные физические механизмы неспецифических эффектов остаются во многом неясными, хотя понимание их природы постепенно углубляется. Знание молекулярных механизмов позволило бы существенно снизить уровень случайности неспецифических эффектов, т.е. повысить их воспроизводимость, и найти им множество полезных применений.
В числе перспективных — механизмы, основанные на квантовой динамике магнитных моментов, — прежде всего магнетохимический механизм, или РПМ. Низкая чувствительность РПМ в области слабых МП компенсируется возможным включением спин-коррелированных радикальных пар в работу биополимерных ферментов, в частности ферментов рибосомальной трансляции. Это позволяет поднять магнитную чувствительность РПМ на два-три порядка за счет статистического усиления вероятности локальных ошибок клеточной трансляции. Тем самым, основная проблема магнитобиологии может считаться решенной, так как появляется непротиворечивое объяснение биологических эффектов слабых МП. Определенным объяснительным потенциалом в отношении эффектов МП обладают и механизмы смешения квантовых уровней магнитных моментов и молекулярного гироскопа.
В настоящей работе перечислены также механизмы, которые в разное время предлагались для объяснения неспецифических биологических эффектов слабых МП, но оказались несостоятельны по тем или иным причинам. К ним относятся индукционные и резонансные механизмы, а также механизмы на основе динамики магнитных наночастиц. Несостоятельны вообще все механизмы, использующие классическую, а не квантовую, физику для описания микроскопических процессов, — например те, что используют представление о силе Лоренца.
Показано, что исследование действия гипоМП на клеточные процессы в условиях компенсации или экранирования лабораторных магнитных флуктуаций имеет хорошие перспективы разнообразных практических применений.
Приложения
1. Механизм смешения квантовых уровней
Помимо специфических эффектов магниторецепции, как сказано выше, имеются и неспецифические эффекты — эпифеномен жизнедеятельности организмов в геоМП. Среди частых свойств неспецифических откликов — это, например, чувствительность к реверсу направления МП и выраженная частотная селективность. РПМ не способен объяснить такие свойства. Кроме того, имеются десятки исследований, в которых наблюдаемые закономерности магнитного отклика также не укладываются в РПМ-сценарий по тем или другим причинам, см. напр. [52,104]. В частности, магнитный отклик наблюдают при возбуждении фотонами красного спектрального диапазона, энергии которых недостаточно для рождения радикальных пар [35], или вообще в темноте.
Для объяснения таких эффектов, по-видимому, нужен какой-то другой механизм. И отдельный МП сенсор должен реагировать на изменение знака МП по крайней мере. Нечувствительность РПМ к реверсу МП обусловлена тем, что МП меняет динамику пары моментов относительно друг друга. Значит, имеет смысл сценарий, где МП меняло бы динамику одиночного момента относительно выделенного направления, заданного локальным молекулярным окружением [105]. Это и есть механизм смешения квантовых уровней [27,62,66].
Сценарий, где МП меняет динамику одиночного момента относительно локального выделенного направления, обладает привлекательными свойствами. Он способен объяснить, пока качественно, особенности неспецифических эффектов и дает простые количественные соотношения для верификации [66]. Механизм является предельно абстрактным и общим, так что альтернатив среди одночастичных квантовых механизмов попросту не может быть. Его выводы не зависят от природы магнитных моментов, и проверяемы в эксперименте.
В применении к спиновым магнитным моментам механизм является квантовым, поскольку спин — это свойство микрочастиц, подчиняющееся законам квантовой физики. В частном случае одноосного МП, остающегося коллинеарным какой-либо оси, допустимо описание механизма при помощи векторной модели спина в рамках классической физики, что представляет значительные математические удобства.
Как сказано выше, параметр управляет вероятностью реакции сенсора на МП. Экспериментально найденное значение МП 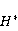 , соответствующее наибольшей чувствительности, связано, следовательно, с произведением , соответствующее наибольшей чувствительности, связано, следовательно, с произведением 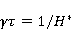 . А это позволяет уточнить природу первичной мишени МП. Действительно, возможных мишеней мало, — электрон, протон, другое магнитное ядро, орбитальный момент электрона или заряженной молекулярной группы, — а значения . А это позволяет уточнить природу первичной мишени МП. Действительно, возможных мишеней мало, — электрон, протон, другое магнитное ядро, орбитальный момент электрона или заряженной молекулярной группы, — а значения 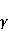 и у них существенно разные и часто известны по порядку величины для разных молекулярных окружений. и у них существенно разные и часто известны по порядку величины для разных молекулярных окружений.
Кроме того, часть первичных сенсоров МП находятся на вращающихся молекулах [62]. Расчеты показывают, что отклик вращающегося магнитного сенсора на МП сдвинут относительно 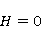 . При этом МП , теперь соответствующее максимуму отклика, связано со скоростью молекулярного вращения . При этом МП , теперь соответствующее максимуму отклика, связано со скоростью молекулярного вращения 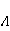 соотношением соотношением 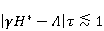 . Видно, что величина эффекта меняется при изменении МП на противоположно направленное, чего нет в РМП, и что позволяет различать молекулярные механизмы экспериментально. Экспериментальное определение зависимости биологического отклика от величины МП способно дать информацию о природе молекулярных процессов неспецифической реакции организмов на слабое МП. В частности, авторы экспериментов [35,106] интерпретируют свои результаты на основе механизма смешения уровней, полагая неудовлетворительным объяснение на основе РПМ. . Видно, что величина эффекта меняется при изменении МП на противоположно направленное, чего нет в РМП, и что позволяет различать молекулярные механизмы экспериментально. Экспериментальное определение зависимости биологического отклика от величины МП способно дать информацию о природе молекулярных процессов неспецифической реакции организмов на слабое МП. В частности, авторы экспериментов [35,106] интерпретируют свои результаты на основе механизма смешения уровней, полагая неудовлетворительным объяснение на основе РПМ.
Описание механизма смешения квантовых уровней требует, в отличие от РПМ, лишь минимального числа переменных и параметров. Недостатком механизма является обратная сторона его предельной абстрактности, общности и универсальности. Механизм ничего не говорит о природе магнитных моментов и способе их связи с биофизическим окружением, хотя и позволяет получить новое знание о природе моментов из сопоставления своих предсказаний с экспериментом.
2. Молекулярный гироскоп
Последний сценарий влияния МП на магнитный момент — это влияние на магнитный момент пространственного вращения заряженных атомно-молекулярных групп, — на орбитальный магнитный момент. Вращение зарядов приводит к возникновению магнитного момента, взаимодействующего с внешним МП. МП изменяет пространственную структуру или фазы угловых квантовых состояний вращающихся молекулярных групп, что приводит к интерференционным эффектам [74] с. 385. Интерференция означает здесь неоднородность плотности вероятности различных угловых позиций молекулярной группы. Магнитное поле, влияя на фазы квантовых состояний, влияет на интерференцию и, тем самым, меняет вероятности пребывания молекулярной группы в различных угловых положениях. Например, изменение вероятности различных угловых позиций аминокислотных остатков в процессе фолдинга белковой цепи могло бы сказываться на скорости или результатах фолдинга.
Вращающаяся молекулярная группа, связанная двумя ковалентными связями, или «опорами», со стенками полости, образованной молекулярным окружением, является молекулярным гироскопом. Короткие отрезки полипептидов или части нуклеиновых кислот могли бы рассматриваться как молекулярные гироскопы, чувствительные к МП. Например уотсон-криковские пары азотистых оснований, скрепляющие две нити ДНК в двойную спираль, обладают вращательной степенью свободы. Их вращения затруднены вследствие стерических ограничений. Однако в области работы ДНК-полимераз стерические ограничения могли бы быть ослаблены и допускать почти свободное вращение пар в некотором интервале времени.
Важно, что тепловые колебания опор молекулярного гироскопа создают лишь нулевые моменты сил относительно его собственной оси вращения. Это означает, что гироскопическая степень свободы релаксирует медленно, — обладает иммунитетом к тепловым колебаниям опор. Оценки термализации за счет ван-дер-ваальсовых взаимодействий, проведенные в [70], показали, что если реалистичный молекулярный гироскоп, образованный аминокислотным остатком фенилаланина, встроен в полость радиуса 1.4 нм, то время релаксации его когерентного вращения составляет 10 мс по порядку величины. Время релаксации растет экспоненциально с размером полости, и при радиусе 1.6 нм достигает 1 с. Гиромагнитное отношение молекулярного гироскопа на 5–6 порядков меньше электронного из-за большой массы гироскопа, но время релаксации когерентности, как видно, на 6–9 порядков больше электронного. Поэтому фундаментальное соотношение , управляющее возникновением магнитных эффектов, могло бы здесь быть выполнено.
Пока неясно, есть ли реально молекулярные конструкции типа гироскопа, обладающие большим временем релаксации. С одной стороны, наличие необходимых для этого и более-менее свободных от воды полостей радиусом около 1.5 нм проблематично. С другой стороны, такие полости не обязательно должны существовать как неизменные структуры. Достаточно, чтобы они появлялись как виртуальные биофизические образования со временем жизни 0.01–1 с. Не исключено, что такие полости могли бы образовываться в ходе фолдинга и созревания длинных белковых молекул, в ходе различных конформационных преобразований хроматин-ДНК комплексов.
Предсказательная сила гироскопического механизма такая же, как и у механизма смешения квантовых уровней. Теоретические оценки показывают богатое особенностями поведение молекулярного гироскопа в переменном МП, в постоянном МП и, в частности, в условиях гипоМП [27]. Недостаток механизма в том, что на сегодня он остается умозрительной концепцией, никак не подтвержденной экспериментально.
3. Несостоятельные механизмы биологических эффектов слабых МП
Ниже перечислены магнитные явления, которые не могут лежать в основе неспецифической чувствительности организмов к МП.
Магнитная индукция
Это индукция вихревых токов высокочастотным или импульсным МП. Обычно вихревые токи импульсных МП значительно превышают токи, создаваемые МП в магнитобиологии неспецифических эффектов. Один из нормативов ICNIRP [20] для населения устанавливает величину 2 мА/м в качестве основного ограничения плотности тока в качестве основного ограничения плотности тока  частотой 4–1000 Гц в организме человека. Можно допустить, что заметные эффекты возникают там, где через организм проходит ток несколько большей плотности. Возьмем для оценок частотой 4–1000 Гц в организме человека. Можно допустить, что заметные эффекты возникают там, где через организм проходит ток несколько большей плотности. Возьмем для оценок 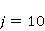 мА/м. Например, в [107] уровень транскрипции в клетках HL-60 изменялся до 30% при плотности тока от 3 до 30 мА/м. Используя закон Ома мА/м. Например, в [107] уровень транскрипции в клетках HL-60 изменялся до 30% при плотности тока от 3 до 30 мА/м. Используя закон Ома 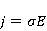 и закон электромагнитной индукции Фарадея и закон электромагнитной индукции Фарадея 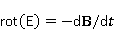 , где , где 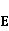 и и 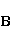 — векторы напряженности электрического поля и плотности магнитного потока, легко оценить скорость изменения МП в сантиметровом образце с проводимостью — векторы напряженности электрического поля и плотности магнитного потока, легко оценить скорость изменения МП в сантиметровом образце с проводимостью 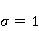 См/м, при которой возникает индукция тока с плотностью мА/м. Скорость составляет 1 Тл/с. Для того чтобы синусоидальное 50-Гц МП влекло такие быстрые изменения, оно должно иметь амплитуду около 20 мТл. Однако многие магнитобиологические эффекты возникают в МП в тысячу раз меньших и даже в постоянных МП, не вызывающих вихревых токов. В этих случаях вихревые токи в теле, как и нагрев, незначительны или невозможны. Таким образом механизмы, основанные на явлении электромагнитной индукции не в состоянии объяснить молекулярные биологические эффекты слабого низкочастотного и, тем более, постоянного МП. См/м, при которой возникает индукция тока с плотностью мА/м. Скорость составляет 1 Тл/с. Для того чтобы синусоидальное 50-Гц МП влекло такие быстрые изменения, оно должно иметь амплитуду около 20 мТл. Однако многие магнитобиологические эффекты возникают в МП в тысячу раз меньших и даже в постоянных МП, не вызывающих вихревых токов. В этих случаях вихревые токи в теле, как и нагрев, незначительны или невозможны. Таким образом механизмы, основанные на явлении электромагнитной индукции не в состоянии объяснить молекулярные биологические эффекты слабого низкочастотного и, тем более, постоянного МП.
Электромагнитную индукцию надо учитывать при интерпретации эффектов в высокочастотном или в импульсном МП. Поскольку вероятность индукционных эффектов пропорциональна частоте МП, наблюдаемый биологический отклик может быть ошибочно приписан магнитному эффекту.
Магнитный резонанс
Магнитный резонанс, как электронный, так и ядерный, не может быть объяснением неспецифических эффектов МП. Магнитный резонанс лежит в основе, например, медицинской томографии и научной магнитной спектрометрии. Для наблюдения магнитного резонанса требуются особые условия: сильное статическое поле в несколько Тл и перпендикулярное ему переменное МП с высокочастотной составляющей, определяемой статическим МП. Условия магнитобиологических эффектов далеки от этих жестких требований. Исключены и другие резонансные явления, где речь идет о накоплении энергии слабого МП в колебаниях атомов и молекул. В низкоинтенсивном МП такое накопление даже в идеализированных условиях потребовало бы когерентности колебаний в течение месяцев и более [22] с. 366, что абсурдно.
В литературе встречается предположение о том, что колебания МП Земли на частотах шумановских резонансов могут вызывать биологический отклик в организме человека вследствие совпадения с частотами 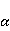 - и других ритмов мозговой деятельности. Поверхность Земли и ионизованный слой атмосферы образуют волновод, в котором распространяются электромагнитные волны. Частота собственных колебаний поля в таком волноводе определяется условием кратности длины электромагнитной волны и длины окружности Земли. Резонансы возбуждаются грозовой деятельностью с интенсивными молниями. В. Шуман рассчитал частоты резонанса с учетом диссипации и сферической симметрии: 8, 14, 20 Гц и т.д. Наиболее выражен резонанс на самой низкой частоте около 8 Гц, соответствующей -ритму. - и других ритмов мозговой деятельности. Поверхность Земли и ионизованный слой атмосферы образуют волновод, в котором распространяются электромагнитные волны. Частота собственных колебаний поля в таком волноводе определяется условием кратности длины электромагнитной волны и длины окружности Земли. Резонансы возбуждаются грозовой деятельностью с интенсивными молниями. В. Шуман рассчитал частоты резонанса с учетом диссипации и сферической симметрии: 8, 14, 20 Гц и т.д. Наиболее выражен резонанс на самой низкой частоте около 8 Гц, соответствующей -ритму.
Резонансные колебания электромагнитного поля могут быть обнаружены на поверхности Земли по измерениям электрической или магнитной компонент. Однако эти колебания не могут вызвать магнитный биологический отклик, поскольку одного только совпадения частот резонанса и -ритма недостаточно. Надо, по крайней мере, чтобы магнитуда колебаний поля была достаточно большой. Величина же шумановских колебаний МП на поверхности Земли ничтожно мала, порядка 1 пТл. МП изменится на эту величину, если, например, включить карманный фонарик в десяти метрах от места измерения. Это на несколько порядков меньше интенсивности городских магнитных флуктуаций тех же частот [22] с. 202. Бесполезность этой концепции видна и в том, что сегодня не ясна природа биологического действия МП порядка 1 мкТл, т.е. полей в миллион раз более сильных, чем шумановские магнитные сигналы.
Магнитные наночастицы
Часто обсуждают возможную вовлеченность магнитных наночастиц в неспецифические магнитные эффекты. Действительно, во многих организмах обнаружены биогенные частицы магнетита размером до десятков нанометров. Их энергия в геоМП часто превышает — характерный масштаб энергии активации химических реакций [108,109], где постоянная Больцмана, — абсолютная температура. В МП наночастицы подвергаются крутящему моменту и могли бы сжимать близлежащие биофизические структуры. Однако в отношении неспецифических эффектов эта идея пока не находит ни теоретического [27], ни экспериментального подтверждения. Кроме того, магнитный эффект наблюдается и в культурах, где магнитные наночастицы заведомо исключают, — в различных бактериях, в культурах клеток животных и растений. По этой причине механизмы, основанные на динамике магнитных наночастиц, не могут лежать в основе неспецифических магнитных эффектов.
Сила Лоренца
Существует определенная закономерность в хаотическом движении свободных ионов в биологических тканях в МП. Под действием силы Лоренца — перпендикулярно вектору скорости и вектору МП — движение иона в промежутках между соударениями с молекулами среды перестает быть прямолинейным. Движение осуществляется по небольшим дугам, так что в среднем возникает общее круговое движение ионов. Такое движение создает очень слабое МП, направленное противоположно приложенному. Другими словами диамагнитный момент среды увеличивается. Однако этот эффект крайне мал [22] стр. 236, на порядки меньше электронного диамагнетизма. Поэтому сила Лоренца невозможна как причина магнитных биологических эффектов в слабом МП.
Библиография
1. von Middendorff, A. T. Die Isepiptesen Russlands. Kaiserlichen Akademie der Wissenschaften, 1855.
2. Schott, H. Zur Geschichte der Elektrotherapie und ihrer Beziehung zum Heilmagnetismus. In Naturheilverfahren und Unkonventionelle Medizinische Richtunge (Ed. V. Fialka und andere) (Springer, 1996).
3. Холодов, Ю. А. Магнетизм в биологии. (Наука, 1970).
4. Холодов, Ю. А. Минуя органы чувств? (Знание, 1991).
5. Пресман, А. С. Действие микроволн на живые организмы и биологические структуры. Успехи Физических Наук 86, 263–302 (1965).
6. Webb, S. J., Dodds, D. E. Microwave inhibition of bacterial growth. Nature 218, 374–375 (1968).
7. Девятков, Н. Д. Влияние электромагнитного излучения миллиметрового диапазона длин волн на биологические объекты. Успехи Физических Наук 110, 453–454 (1973).
8. Тамбиев, А. Х., Кирикова, Н. Н., Бецкий, О. В., Гуляев, Ю. В. Миллиметровые волны и фотосинтезирующие организмы. (Радиотехника, 2003).
9. Belpomme, D., Hardell, L., Belyaev, I., Burgio, E., Carpenter, D. O. Thermal and non-thermal health effects of low intensity non-ionizing radiation: An international perspective. Environmental Pollution 242, 643–658 (2018).
10. Чуян, Е. Н., Джелдубаева, Э. Р. Низкоинтенсивное миллиметровое излучение: Нейроиммуноэндокринные механизмы адаптационных реакций. (ИТ АРИАЛ, 2020).
11. Пресман, А. С. Электромагнитные поля и живая природа. (Hаука, 1968).
12. Аникин, В. М. Магнитобиология и магнитотерапия: Кармиловский период. Гетеромагнитная Микроэлектроника 131–140 (2016).
13. Moulder, J. E. The electric and magnetic fields research and public information dissemination (EMF-RAPID) program. Radiation Research 153, 613–616 (2000).
14. IARC. IARC monographs on the evaluation of carcinogenic risks to humans, vol. 80. Non-ionizing radiation, part 1: Static and extremely low-frequency (ELF) electric and magnetic fields. (IARC Press, 2002).
15. Huss, A., Peters, S., Vermeulen, R. Occupational exposure to extremely low-frequency magnetic fields and the risk of ALS: A systematic review and meta-analysis. Bioelectromagnetics 39, 156–163 (2018).
16. Miller, A. B., Morgan, L. L., Udasin, I., Davis, D. L. Cancer epidemiology update, following the 2011 IARC evaluation of radiofrequency electromagnetic fields (Monograph 102). Environmental Research 167, 673–683 (2018).
17. Li, D.-K., Chen, H., Ferber, J. R., Odouli, R., Quesenberry, Ch. Exposure to magnetic field non-ionizing radiation and the risk of miscarriage: A prospective cohort study. Scientific Reports 7, 17541 (2017).
18. Belyaev, I., Dean, A., Eger, H., Hubmann, G., Jandrisovits, R., Kern, M., et al. EUROPAEM EMF Guideline 2016 for the prevention, diagnosis and treatment of EMF-related health problems and illnesses. Reviews on Environmental Health 31, 363–397 (2016).
19. Environmental health criteria 238. Extremely low frequency fields. (World Health Organization, 2007).
20. International Commission on Non-Ionizing Radiation Protection. Guidelines for limiting exposure to time-varying, electric, magnetic and electromagnetic fields up to 300 GHz. Health Physics 74, 494–521 (1998).
21. Wiltschko, W., Wiltschko, R. Magnetic compass of european robins. Science 176, 62–64 (1972).
22. Бинги, В. Н. Принципы электромагнитной биофизики. (Физматлит, 2011).
23. Roosli, M., ed. Epidemiology of Electromagnetic Fields. (CRC Press, 2014).
24. Кудряшов, Ю. Б., Рубин, А. Б. Радиационная биофизика: Сверхнизкочастотные электромагнитные излучения. (Физматлит, 2014).
25. Markov, M. S., ed. Electromagnetic Fields in Biology and Medicine. (CRC Press, 2015).
26. Greenebaum, B., Barnes, F., eds. Biological and Medical Aspects of Electromagnetic Fields. Vols. 1, 2, (CRC Press, 2019).
27. Binhi, V. N., Prato, F. S. Biological effects of the hypomagnetic field: An analytical review of experiments and theories. PLoS ONE 12, e0179340 (2017).
28. Binhi, V. N. Nonspecific magnetic biological effects: A model assuming the spin-orbit coupling. The Journal of Chemical Physics 151, 204101 (2019).
29. Lohmann, K. J., Lohmann, C. M., Ehrhart, L. M., Bagley, D. A., Swing, T. Animal behaviour: Geomagnetic map used in sea-turtle navigation. Nature 428, 909–910 (2004).
30. Bianco, G., Ilieva, M., Åkesson, S. Magnetic storms disrupt nocturnal migratory activity in songbirds. Biology Letters 15, 20180918 (2019).
31. Кишкинев, Д. А., Чернецов, Н. С. Магниторецепторные системы у птиц: Обзор современных исследований. Журнал Общей Биологии 75, 104–123 (2014).
32. Hore, P. J., Mouritsen, H. The radical-pair mechanism of magnetoreception. Annual Review of Biophysics 45, 299–344 (2016).
33. Wan, G., Hayden, A. N., Iiams, S. E., Merlin, C. Cryptochrome 1 mediates light-dependent inclination magnetosensing in monarch butterflies. Nature Communications 12, 771 (2021).
34. Новицкий, Ю. И., Новицкая, Г. В. Действие постоянного магнитного поля на растения. (Наука, 2016).
35. Dhiman, S. K., Wu, F., Galland, P. Effects of weak static magnetic fields on the development of seedlings of arabidopsis thaliana. Protoplasma Sep 21, (2022).
36. Бреус, Т. К., Бинги, В. Н., Петрукович, А. А. Магнитный фактор солнечно-земных связей и его влияние на человека: Физические проблемы и перспективы. Успехи Физических Наук 186, 568–576 (2016).
37. Fu, J.-P., Mo, W.-C., Liu, Y., He, R.-Q. Decline of cell viability and mitochondrial activity in mouse skeletal muscle cell in a hypomagnetic field. Bioelectromagnetics 37, 212–222 (2016).
38. Paponov, I. A., Fliegmann, J., Narayana, R., E., M. M. Differential root and shoot magnetoresponses in arabidopsis thaliana. Scientific Reports 11, 1–15 (2021).
39. Johnsen, S., Lohmann, K. J. The physics and neurobiology of magnetoreception. Nature Reviews. Neuroscience 6, 703–712 (2005).
40. Бучаченко, А. Л. Магнито-зависимые молекулярные и химические процессы в биохимии, генетике и медицине. Успехи Химии 83, 1–12 (2014).
41. Yang, X., Li, Zh., Polyakova, T., Dejneka, A., Zablotskii, V., Zhang, X. Effect of static magnetic field on DNA synthesis: The interplay between DNA chirality and magnetic field left–right asymmetry. FASEB BioAdvances 2, 254–263 (2020).
42. Шредингер, Э. Что такое жизнь? Физический аспект живой клетки. (РХД, 2002).
43. Бинги, В. Н. Физические эффекты сознания: Закон воспроизводимости. pp. 559 (Инфра-М, www.lawr.info, 2021).
44. Иваницкий, Г. Р. XXI век: Что такое жизнь с точки зрения физики. Успехи Физических Наук 180, 337–369 (2010).
45. Marais, A., Adams, B., Ringsmuth, A. K., Ferretti, M., Gruber, J. M., Hendrikx, R., et al. The future of quantum biology. Journal of The Royal Society Interface 15, 20180640 (2018).
46. Сюракшин, А. В., Салеев, В. А., Юшанхай, В. Ю. Квантовые модели в биологии. Вестник Самарского университета. Естественнонаучная серия 28, 74–94 (2022).
47. Binhi, V. N., Rubin, A. B. Theoretical concepts in magnetobiology after 40 years of research. Cells 11, 274 (2022).
48. Sarimov, R. M., Binhi, V. N. Low-frequency magnetic fields in cars and office premises and the geomagnetic field variations. Bioelectromagnetics 41, 360–368 (2020).
49. Makinistian, L., Muehsam, D. J., Bersani, F., Belyaev, I. Some recommendations for experimental work in magnetobiology, revisited. Bioelectromagnetics 39, 556–564 (2018).
50. Prato, F. S., Desjardins-Holmes, D., Keenliside, L. D., DeMoor, J. M., Robertson, J. A., Thomas, A. W. Magnetoreception in laboratory mice: Sensitivity to extremely low frequency fields exceeds 33 nT at 30 Hz. Journal of The Royal Society Interface 10, (2013).
51. Engels, S., Schneider, N.-L., Lefeldt, N., Hein, C. M., Zapka, M., Michalik, A., et al. Anthropogenic electromagnetic noise disrupts magnetic compass orientation in a migratory bird. Nature 509, 353–356 (2014).
52. Bojarinova, J., Kavokin, K., Pakhomov, A., Cherbunin, R., Anashina, A., Erokhina, M., et al. Magnetic compass of garden warblers is not affected by oscillating magnetic fields applied to their eyes. Scientific Reports 10, 3473 (2020).
53. Pishchalnikov, Y., R., Gurfinkel, Yu. I., Sarimov, R. M., Vasin, A. L., Sasonko, M. L., Matveeva, T. A., et al. Cardiovascular response as a marker of environmental stress caused by variations in geomagnetic field and local weather. Biomedical Signal Processing and Control 51, 401–410 (2019).
54. Krylov, V. V., Kantserova, N. P., Lysenko, L. A., Osipova, E. A. A simulated geomagnetic storm unsynchronizes with diurnal geomagnetic variation affecting calpain activity in roach and great pond snail. International Journal of Biometeorology 63, 241–246 (2019).
55. Janashia, K., Tvildiani, L., Tsibadze, T., Invia, N. Effects of the geomagnetic field time-varying components compensation as evidenced by heart rate variability of healthy males. Life Sciences in Space Research 32, 38–44 (2022).
56. Binhi, V. N. Random effects in magnetobiology and a way to summarize them. Bioelectromagnetics 42, 501–515 (2021).
57. Зенченко, Т. А., Бреус, Т. К. Возможные причины нестабильности воспроизведения гелиобиологических результатов. Физика Биологии и Медицины (2023).
58. Valberg, P. A. Designing EMF experiments: What is required to characterize ‘exposure’? Bioelectromagnetics 16, 396–401 (1995).
59. Buchachenko, A. Why magnetic and electromagnetic effects in biology are irreproducible and contradictory? Bioelectromagnetics 37, 1–13 (2016).
60. Portelli, L. Overcoming the irreproducibility barrier. in Bioengineering and biophysical aspects of electromagnetic fields (eds. Greenebaum, B., Barnes, F.) pp. 435–462 (CRC Press, 2019).
61. Koonin, E. V. The Logic of Chance: The Nature and Origin of Biological Evolution. (FT Press, 2012).
62. Binhi, V. N., Prato, F. S. Rotations of macromolecules affect nonspecific biological responses to magnetic fields. Scientific Reports 8, 13495 (2018).
63. Zenchenko, T. A., Breus, T. K. The possible effect of space weather factors on various physiological systems of the human organism. Atmosphere 12, 1–28 (2021).
64. Krylov, V. V., Bolotovskaya, I. V., Osipova, E. A. The response of European Daphnia magna Straus and Australian Daphnia carinata King to changes in geomagnetic field. Electromagnetic Biology and Medicine 32, 30–39 (2013).
65. Васин, А. Л., Шафиркин, А. В., Гурфинкель, Ю. И. Влияние искусственного периодического геомагнитного поля миллигерцового диапазона на показатели вариабельности сердечного ритма. Авиакосмическая и Экологическая Медицина 53, 62–69 (2019).
66. Binhi, V. N., Prato, F. S. A physical mechanism of magnetoreception: extension and analysis. Bioelectromagnetics 38, 41–52 (2017).
67. Buchachenko, A. Magneto-Biology and Medicine. (Nova Science, 2014).
68. Бинги, В. H. Ядерные спины в первичных механизмах биологического действия магнитных полей. Биофизика 40, 677–691 (1995).
69. Бинги, В. Н. Дефекты структуры жидкой воды в магнитном и электрическом полях. Биомедицинская Радиоэлектроника 7–16 (1998).
70. Binhi, V. N., Savin, A. V. Molecular gyroscopes and biological effects of weak extremely low-frequency magnetic fields. Physical Review E 65, 051912 (2002).
71. Fesenko, E. E., Gluvstein, A. Ya. Changes in the state of water, induced by radiofrequency electromagnetic fields. FEBS Lett. 367, 53–55 (1995).
72. Lobyshev, V. I. Water is a sensor to weak forces including electromagnetic fields of low intensity. Electromagnetic Biology and Medicine 24, 449–461 (2005).
73. Бинги, В. Н., Рубин, А. Б. Фундаментальная проблема магнитобиологии. Биомедицинские Технологии и Радиоэлектроника 63–76 (2007).
74. Binhi, V. N. Magnetobiology: Underlying Physical Problems. (Academic Press, 2002).
75. Afanasyeva, M. S., Taraban, M. B., Purtov, P. A., Leshina, T. V., Grissom, C. B. Magnetic spin effects in enzymatic reactions: Radical oxidation of NADH by horseradish peroxidase. Journal of the American Chemical Society 128, 8651–8658 (2006).
76. Buchachenko, A. L., Kuznetsov, D. A., Breslavskaya, N. N. Chemistry of enzymatic ATP synthesis: An insight through the isotope window. Chemical Reviews 112, 2042–2058 (2012).
77. Минаев, Б. Ф. Электронные механизмы активации молекулярного кислорода. Успехи Химии 76, 1059–1083 (2007).
78. Amonkosolpan, J., Aliev, G. N., Wolverson, D., Snow, P. A., Davies, J. J. Magnetic field dependence of singlet oxygen generation by nanoporous silicon. Nanoscale Research Letters 9, 342 (2014).
79. Schulten, K., Swenberg, C., Weller, A. A biomagnetic sensory mechanism based on magnetic field modulated coherent electron spin motion. Zeitschrift Fur Physikalische Chemie 111, 1–5 (1978).
80. Xu, J., Jarocha, L. E., Zollitsch, T., Konowalczyk, M., Henbest, K. B., Richert, S., et al. Magnetic sensitivity of cryptochrome 4 from a migratory songbird. Nature 594, 535–540 (2021).
81. Mouritsen, H. Long-distance navigation and magnetoreception in migratory animals. Nature 558, 50–59 (2018).
82. Hoff, A. J., Rademaker, H., Van Grondelle, R., Duysens, L. N. M. On the magnetic field dependence of the yield of the triplet state in reaction centers of photosynthetic bacteria. Biochimica Et Biophysica Acta – Bioenergetics 460, 547–554 (1977).
83. Steiner, U. E., Ulrich, T. Magnetic field effects in chemical kinetics and related phenomena. Chemical Reviews 89, 51–147 (1989).
84. Fedin, M. V., Shakirov, S. R., Purtov, P. A., Bagryanskaya, E. G. Electron spin relaxation of radicals in weak magnetic fields. Russian Chemical Bulletin 55, 1703–1716 (2006).
85. Ivanov, K. L., Petrova, M. V., Lukzen, N. N., Maeda, K. Consistent treatment of spin-selective recombination of a radical pair confirms the Haberkorn approach. Journal of Physical Chemistry A 114, 9447–9455 (2010).
86. Gauger, E. M., Rieper, E., Morton, J. J. L., Benjamin, S. C., Vedral, V. Sustained quantum coherence and entanglement in the avian compass. Physical Review Letters 106, 040503 (2011).
87. Kattnig, D. R., Solov’yov, I., Hore, P. J. Electron spin relaxation in cryptochrome-based magnetoreception. Physical Chemistry Chemical Physics 18, 12443–12456 (2016).
88. Worster, S., Kattnig, D. R., Hore, P. J. Spin relaxation of radicals in cryptochrome and its role in avian magnetoreception. The Journal of Chemical Physics 145, 035104 (2016).
89. Cai, J., Plenio, M. B. Chemical compass model for avian magnetoreception as a quantum coherent device. Physical Review Letters 111, 230503 (2013).
90. Weaver, J. C., Vaughan, T. E., Astumian, R. D. Biological sensing of small field differences by magnetically sensitive chemical reactions. Nature 405, 707–709 (2000).
91. Grundler, W., Kaiser, F., Keilmann, F., Walleczek, J. Mechanisms of electromagnetic interaction with cellular systems. Naturwissenschaften 79, 551–559 (1992).
92. Riznichenko, G. Yu., Plyusnina, T. Yu., Aksyonov, S. I. Modelling of the effect of a weak electric field on a nonlinear transmembrane ion transfer system. Bioelectrochem. Bioenerg. 35, 39–47 (1994).
93. Player, T. C., Baxter, E. D. A., Allatt, S., Hore, P. J. Amplification of weak magnetic field effects on oscillating reactions. Scientific Reports 11, 9615 (2021).
94. Бинги, В. Н. Магнитная навигация животных, контрастная чувствительность зрения и закон Вебера-Фехнера. Сенсорные Системы 37, 46–60 (2023).
95. Binhi, V. N. Statistical amplification of the effects of weak MFs in cellular translation. Cells 12, 724 (2023).
96. Mohler, K., Ibba, M. Translational fidelity and mistranslation in the cellular response to stress. Nature Microbiology 2, 17117 (2017).
97. Parker, J. Errors and alternatives in reading the universal genetic code. Microbiological Reviews 53, 273–298 (1989).
98. Kurland, C. G. Translational accuracy and the fitness of bacteria. Annual Review of Genetics 26, 29–50 (1992).
99. Nissley, D. A., Jiang, Y., Trovato, F., Sitarik, I., Narayan, K. B., To, P., et al. Universal protein misfolding intermediates can bypass the proteostasis network and remain soluble and less functional. Nature Communications 13, 3081 (2022).
100. Belyaev, I. Y., Koch, C. B., Terenius, O., Roxstrom-Lindquist, K., Malmgren, L. O. G., Sommer, W. H., et al. Exposure of rat brain to 915 MHz GSM microwaves induces changes in gene expression but not double stranded DNA breaks or effects on chromatin conformation. Bioelectromagnetics 27, 295–306 (2006).
101. Drummond, D. A., Wilke, C. O. Mistranslation-induced protein misfolding as a dominant constraint on coding-sequence evolution. Cell 134, 341–352 (2008).
102. Legrain, P., Aebersold, R., Archakov, A., Bairoch, A., Bala, K., Beretta, L., et al. The human proteome project: Current state and future direction. Molecular & Cellular Proteomics 10, (2011).
103. Stovbun, S. V., Zlenko, D. V., Bukhvostov, A. A., Vedenkin, A. A., Skoblin, A. A., Kuznetsov, D. A., et al. Magnetic field and nuclear spin influence on the DNA synthesis rate. Scientific Reports 13, 465 (2023).
104. Agliassa, C., Maffei, M. E. Reduction of geomagnetic field (GMF) to near null magnetic field (NNMF) affects some Arabidopsis thaliana clock genes amplitude in a light independent manner. Journal of Plant Physiology 232, 23–26 (2019).
105. Бинги, В. Н. Первичный физический механизм биологических эффектов слабых магнитных полей. Биофизика 61, 201–208 (2016).
106. Dhiman, S. K., Galland, P. Effects of weak static magnetic fields on the gene expression of seedlings of Arabidopsis thaliana. Journal of Plant Physiology 231, 9–18 (2018).
107. Blank, M., Soo, L., Lin, H., Henderson, A. S., Goodman, R. Changes in transcription in HL-60 cells following exposure to alternating currents from electric fields. Bioelectrochemistry and Bioenergetics 28, 301–309 (1992).
108. Binhi, V. N., Chernavsky, D. S. Stochastic resonance of magnetosomes fixed in the cytoskeleton. Biophysics 50, 599–603 (2005).
109. Kirschvink, J. L., Winklhofer, M., Walker, M. M. Biophysics of magnetic orientation: Strengthening the interface between theory and experimental design. Journal of the Royal Society Interface 7, S179–S171 (2010).
References
1. Middendorff, A. T. Die Isepiptesen Russlands. (1855). Kaiserlichen Akademie der Wissenschaften.
2. Schott, H. Zur. (1996). Geschichte der Elektrotherapie und ihrer Beziehung zum Heilmagnetismus. In Naturheilverfahren und Unkonventionelle Medizinische Richtunge (Ed. V. Fialka und andere). Springer.
3. Kholodov, Yu. A. (1970). Magnetism in Biology. Nauka.
4. Kholodov, Yu. A. (1991). Bypassing the Sensory Organs? Znanie.
5. Presman, A. S. (1965). The action of microwaves on living organisms and biological structures. Physics-Uspekhi 8, 463–488.
6. Webb, S. J., Dodds, D. E. (1968). Microwave inhibition of bacterial growth. Nature 218, 374–375.
7. Devyatkov, N. D. (1974). Influence of millimeter-band electromagnetic radiation on biological objects. Physics–Uspekhi 16, 568–569.
8. Tambiyev, A. H., & Kirikova, & N. N., & Betskii, & O. V., Gulyaev, Y. V. (2003). Millimeter Waves and Photosynthetic Organisms. Radioengineering.
9. Belpomme, D., & Hardell, L., & Belyaev, & I., & Burgio, E., & Carpenter, D. O. (2018). Thermal and non-thermal health effects of low intensity non-ionizing radiation: An international perspective. Environmental Pollution 242, 643–658.
10. Chuyan, E. N., & Djeldubaeva, E. R. (2020). Low-Intensity Millimeter-Wave Radiation: Neuro Immunoendocrine Mechanisms of Adaptative Reactions. IT ARIAL.
11. Presman, A. S. (1968). Electromagnetic Fields and Wildlife. Nauka.
12. Anikin, V. M. (2016). Magnetobiology and magnetotherapy: Karmilov period. Heteromagnetic Microelectronics 21, 131–140.
13. Moulder, J. E. (2000). The electric and magnetic fields research and public information dissemination (EMF-RAPID) program. Radiation Research 153, 613–616.
14. IARC. (2002). IARC monographs on the evaluation of carcinogenic risks to humans. Vol. 80. Non-ionizing radiation, part 1: Static and extremely low-frequency (ELF) electric and magnetic fields. IARC Press.
15. Huss, A., & Peters, S., & Vermeulen, R. (2018). Occupational exposure to extremely low-frequency magnetic fields and the risk of ALS: A systematic review and meta-analysis. Bioelectromagnetics 39, 156–163.
16. Miller, A. B., & Morgan, L. L., & Udasin, I., & Davis, D. L. (2018). Cancer epidemiology update, following the 2011 IARC evaluation of radiofrequency electromagnetic fields (Monograph 102). Environmental Research, 167, 673–683.
17. Li, D.-K., & Chen, H., & Ferber, J. R., & Odouli, R., & Quesenberry, Ch. (2017). Exposure to magnetic field non-ionizing radiation and the risk of miscarriage: A prospective cohort study. Scientific Reports 7, 17541.
18. Belyaev, I., & Dean, A., & Eger, H., & Hubmann, & G., Jandrisovits, R., & Kern, M., et al. (2016). EUROPAEM EMF Guideline 2016 for the prevention, diagnosis and treatment of EMF-related health problems and illnesses. Reviews on Environmental Health, 31, 363–397.
19. Environmental health criteria 238. (2007). Extremely low frequency fields. World Health Organization.
20. International Commission on Non-Ionizing Radiation Protection. (1998). Guidelines for limiting exposure to time-varying, electric, magnetic and electromagnetic fields up to 300 GHz. Health Physics 74, 494–521.
21. Wiltschko, W., & Wiltschko, R. (1972). Magnetic compass of european robins. Science, 176, 62–64.
22. Binhi, V. N. (2011). Principles of Electromagnetic Biophysics. Fizmatlit.
23. Roosli, M. (Ed.). (2014). Epidemiology of Electromagnetic Fields. CRC Press.
24. Kudryashov, Yu. B., & Rubin, A. B. (2014). Radiation Biophysics: Ultra-Low Frequency Electromagnetic Radiation. Fizmatlit.
25. Markov, M. S. (Ed.). (2015). Electromagnetic Fields in Biology and Medicine. CRC Press.
26. Greenebaum, B., & Barnes, F. (2019) Biological and Medical Aspects of Electromagnetic Fields. Vols. 1, 2. CRC Press.
27. Binhi, V. N., & Prato, F. S. (2017). Biological effects of the hypomagnetic field: An analytical review of experiments and theories. PLoS ONE, 12, e0179340.
28. Binhi, V. N. (2019). Nonspecific magnetic biological effects: A model assuming the spin-orbit coupling. The Journal of Chemical Physics, 151, 204101.
29. Lohmann, K. J., & Lohmann, C. M., & Ehrhart, L. M., & Bagley, D. A., & Swing, T. (2004). Animal behaviour: Geomagnetic map used in sea-turtle navigation. Nature, 428, 909–910.
30. Bianco, G., Ilieva, M., Åkesson, S. (2019). Magnetic storms disrupt nocturnal migratory activity in songbirds. Biology Letters, 15, 20180918.
31. Kishkinev, D. A., & Chernetsov, N. S. (2014). Magnetoreception systems in birds: A review of current research. General Biology Journal, 75, 104–123.
32. Hore, P. J., & Mouritsen, H. The radical-pair mechanism of magnetoreception. Annual Review of Biophysics, 45, 299–344 (2016).
33. Wan, G., & Hayden, A. N., Iiams, & S. E., Merlin, C. (2021). Cryptochrome 1 mediates light-dependent inclination magnetosensing in monarch butterflies. Nature Communications, 12, 771.
34. Novitsky, Yu. I., & Novitskaya, G. V. (2016). The Effect of a Constant Magnetic Field on Plants. Nauka.
35. Dhiman, S. K., & Wu, F., & Galland, P. (2022). Effects of weak static magnetic fields on the development of seedlings of arabidopsis thaliana. Protoplasma Sep 21.
36. Breus, T. K., & Binhi, V. N., & Petrukovich, A. A. (2016). Magnetic factor of the solar terrestrial relations and its impact on the human body: physical problems and prospects for research. Physics–Uspekhi, 59, 502–510.
37. Fu, J.-P., & Mo, W.-C., & Liu, Y., & He, R.-Q. (2016). Decline of cell viability and mitochondrial activity in mouse skeletal muscle cell in a hypomagnetic field. Bioelectromagnetics, 37, 212–222.
38. Paponov, I. A., & Fliegmann, J., & Narayana, R., & E., M. M. (2021). Differential root and shoot magnetoresponses in arabidopsis thaliana. Scientific Reports, 11, 1–15.
39. Johnsen, S., & Lohmann, K. J. (2005). The physics and neurobiology of magnetoreception. Nature Reviews. Neuroscience, 6, 703–712.
40. Buchachenko, A. L. (2014). Magnetic field-dependent molecular and chemical processes in biochemistry, genetics and medicine. Russian Chemical Reviews, 83, 1–12.
41. Yang, X., & Li, Zh., Polyakova, T., & Dejneka, A., & Zablotskii, V., & Zhang, X. (2020). Effect of static magnetic field on DNA synthesis: The interplay between DNA chirality and magnetic field left–right asymmetry. FASEB BioAdvances, 2, 254–263.
42. Schrödinger, E. (2002). What is Life? The Physical Aspect of the Living Cell. R&C Dynamics.
43. Binhi, V. N. (2021). Physical Effects of Consciousness: The Law of Reproducibility. Infra-M, www.lawr.info.
44. Ivanitskii, G. R. (2010). 21st century: What is life from the perspective of physics? Physics-Uspekhi, 53, 327–356.
45. Marais, A., & Adams, B., & Ringsmuth, A. K., & Ferretti, M., & Gruber, J. M., & Hendrikx, R., et al. (2018). The future of quantum biology. Journal of The Royal Society Interface, 15, 20180640.
46. Syurakshin, A. V., & Saleev, V. A., & Yushankhai, V. Yu. (2022). Quantum models in biology. Vestnik of Samara University. Natural Science Series, 28, 74–94.
47. Binhi, V. N., & Rubin, A. B. (2022). Theoretical concepts in magnetobiology after 40 years of research. Cells, 11, 274.
48. Sarimov, R. M., Binhi, V. N. (2020). Low-frequency magnetic fields in cars and office premises and the geomagnetic field variations. Bioelectromagnetics, 41, 360–368.
49. Makinistian, L., & Muehsam, D. J., & Bersani, F., & Belyaev, I. (2018). Some recommendations for experimental work in magnetobiology, revisited. Bioelectromagnetics, 39, 556–564.
50. Prato, F. S., & Desjardins-Holmes, D., & Keenliside, L. D., & DeMoor, J. M., & Robertson, J. A., & Thomas, A. W. (2013). Magnetoreception in laboratory mice: Sensitivity to extremely low frequency fields exceeds 33 nT at 30 Hz. Journal of The Royal Society Interface, 10.
51. Engels, S., & Schneider, N.-L., & Lefeldt, N., & Hein, C. M., & Zapka, M., & Michalik, A., et al. (2014). Anthropogenic electromagnetic noise disrupts magnetic compass orientation in a migratory bird. Nature, 509, 353–356.
52. Bojarinova, J., & Kavokin, K., & Pakhomov, A., & Cherbunin, R., & Anashina, A., & Erokhina, M., et al. (2020). Magnetic compass of garden warblers is not affected by oscillating magnetic fields applied to their eyes. Scientific Reports, 10, 3473.
53. Pishchalnikov, Y., R., & Gurfinkel, Yu. I., & Sarimov, R. M., & Vasin, A. L., & Sasonko, M. L., & Matveeva, T. A., et al. (2019). Cardiovascular response as a marker of environmental stress caused by variations in geomagnetic field and local weather. Biomedical Signal Processing and Control, 51, 401–410.
54. Krylov, V. V., & Kantserova, N. P., & Lysenko, L. A., & Osipova, E. A. (2019). A simulated geomagnetic storm unsynchronizes with diurnal geomagnetic variation affecting calpain activity in roach and great pond snail. International Journal of Biometeorology, 63, 241–246.
55. Janashia, K., & Tvildiani, L., & Tsibadze, T., & Invia, N. (2022). Effects of the geomagnetic field time-varying components compensation as evidenced by heart rate variability of healthy males. Life Sciences in Space Research, 32, 38–44.
56. Binhi, V. N. (2021). Random effects in magnetobiology and a way to summarize them. Bioelectromagnetics, 42, 501–515.
57. Zenchenko, T. A., & Breus, T. K. (2023). Possible causes for the instability of reproduction of heliobiological results. Physics of Biology and Medicine.
58. Valberg, P. A. (1995). Designing EMF experiments: What is required to characterize ‘exposure’? Bioelectromagnetics, 16, 396–401.
59. Buchachenko, A. (2016). Why magnetic and electromagnetic effects in biology are irreproducible and contradictory? Bioelectromagnetics, 37, 1–13.
60. Portelli, L. (2019). Overcoming the irreproducibility barrier. in Bioengineering and biophysical aspects of electromagnetic fields (eds. Greenebaum, B., Barnes, F.) pp. 435–462. CRC Press.
61. Koonin, E. V. (2012). The Logic of Chance: The Nature and Origin of Biological Evolution. FT Press.
62. Binhi, V. N., & Prato, F. S. (2018). Rotations of macromolecules affect nonspecific biological responses to magnetic fields. Scientific Reports, 8, 13495.
63. Zenchenko, T. A., & Breus, T. K. (2021). The possible effect of space weather factors on various physiological systems of the human organism. Atmosphere, 12, 1–28.
64. Krylov, V. V., & Bolotovskaya, & I. V., Osipova, & E. A. (2013). The response of European Daphnia magna Straus and Australian Daphnia carinata King to changes in geomagnetic field. Electromagnetic Biology and Medicine, 32, 30–39.
65. Vasin, A. L., & Shafirkin, A. V., & I., G. Yu. (2019). Effect of artificial alternating geomagnetic field in the millihertz range on the heart rate variability indices. Aerospace and Environmental Medicine, 53, 62–69.
66. Binhi, V. N., & Prato, F. S. (2017). A physical mechanism of magnetoreception: extension and analysis. Bioelectromagnetics, 38, 41–52.
67. Buchachenko, A. (2014). Magneto-Biology and Medicine. Nova Science.
68. Binhi, V. N. 1995. Nuclear spins in the primary mechanisms of biological action of magnetic fields. Biophysics, 40, 671–685.
69. Binhi, V. N. (1998). On structural defects of liquid water in magnetic and electric fields. Biomedical Radioelektronics, 7, 16.
70. Binhi, V. N., & Savin, A. V. (2002). Molecular gyroscopes and biological effects of weak extremely low-frequency magnetic fields. Physical Review, E 65, 051912.
71. Fesenko, E. E., & Gluvstein, A. Ya. (1995). Changes in the state of water, induced by radiofrequency electromagnetic fields. FEBS Lett., 367, 53–55.
72. Lobyshev, V. I. (2005). Water is a sensor to weak forces including electromagnetic fields of low intensity. Electromagnetic Biology and Medicine, 24, 449–461.
73. Binhi, V. N., & Rubin, A. B. (2007). Fundamental problem of magnetobiology. Biomedical Technologies and Radioelectronics, 63, 76.
74. Binhi, V. N. (2002). Magnetobiology: Underlying Physical Problems. Academic Press.
75. Afanasyeva, M. S., & Taraban, M. B., & Purtov, P. A., & Leshina, T. V., & Grissom, C. B. (2006). Magnetic spin effects in enzymatic reactions: Radical oxidation of NADH by horseradish peroxidase. Journal of the American Chemical Society, 128, 8651–8658.
76. Buchachenko, A. L., & Kuznetsov, D. A., & Breslavskaya, N. N. (2012). Chemistry of enzymatic ATP synthesis: An insight through the isotope window. Chemical Reviews, 112, 2042–2058.
77. Minaev, B. F. (2007). Electronic mechanisms of molecular oxygen activation. Russian Chemical Reviews, 76, 989–1011.
78. Amonkosolpan, J., & Aliev, G. N., & Wolverson, D., & Snow, P. A., & Davies, J. J. (2014). Magnetic field dependence of singlet oxygen generation by nanoporous silicon. Nanoscale Research Letters, 9, 342.
79. Schulten, K., & Swenberg, C., & Weller, A. (1978). A biomagnetic sensory mechanism based on magnetic field modulated coherent electron spin motion. Zeitschrift Fur Physikalische Chemie, 111, 1–5.
80. Xu, J., & Jarocha, L. E., & Zollitsch, T., & Konowalczyk, M., & Henbest, K. B., & Richert, S., et al. (2021). Magnetic sensitivity of cryptochrome 4 from a migratory songbird. Nature, 594, 535–540.
81. Mouritsen, H. (2018). Long-distance navigation and magnetoreception in migratory animals. Nature, 558, 50–59.
82. Hoff, A. J., & Rademaker, H., & Van Grondelle, & R., Duysens, L. N. M. (1977). On the magnetic field dependence of the yield of the triplet state in reaction centers of photosynthetic bacteria. Biochimica Et Biophysica Acta – Bioenergetics, 460, 547–554.
83. Steiner, U. E., & Ulrich, T. (1989). Magnetic field effects in chemical kinetics and related phenomena. Chemical Reviews, 89, 51–147.
84. Fedin, M. V., & Shakirov, S. R., & Purtov, P. A., & Bagryanskaya, E. G. (2006). Electron spin relaxation of radicals in weak magnetic fields. Russian Chemical Bulletin, 55, 1703–1716.
85. Ivanov, K. L., & Petrova, M. V., & Lukzen, N. N., & Maeda, K. (2010). Consistent treatment of spin-selective recombination of a radical pair confirms the Haberkorn approach. Journal of Physical Chemistry A, 114, 9447–9455.
86. Gauger, E. M., & Rieper, E., & Morton, J. J. L., & Benjamin, S. C., & Vedral, V. (2011). Sustained quantum coherence and entanglement in the avian compass. Physical Review Letters, 106, 040503.
87. Kattnig, D. R., & Solov’yov, I., & Hore, P. J. (2016). Electron spin relaxation in cryptochrome-based magnetoreception. Physical Chemistry Chemical Physics, 18, 12443–12456.
88. Worster, S., & Kattnig, D. R., & Hore, P. J. (2016). Spin relaxation of radicals in cryptochrome and its role in avian magnetoreception. The Journal of Chemical Physics, 145, 035104.
89. Cai, J., & Plenio, M. B. (2013). Chemical compass model for avian magnetoreception as a quantum coherent device. Physical Review Letters, 111, 230503.
90. Weaver, J. C., & Vaughan, T. E., & Astumian, R. D. (2000). Biological sensing of small field differences by magnetically sensitive chemical reactions. Nature, 405, 707–709.
91. Grundler, W., & Kaiser, F., & Keilmann, F., & Walleczek, J. (1992). Mechanisms of electromagnetic interaction with cellular systems. Naturwissenschaften, 79, 551–559.
92. Riznichenko, G. Yu., & Plyusnina, T. Yu., & Aksyonov, S. I. (1994). Modelling of the effect of a weak electric field on a nonlinear transmembrane ion transfer system. Bioelectrochem. Bioenerg., 35, 39–47.
93. Player, T. C., & Baxter, E. D. A., & Allatt, S., & Hore, P. J. (2021). Amplification of weak magnetic field effects on oscillating reactions. Scientific Reports, 11, 9615.
94. Binhi, V. N. (2023). Magnetic navigation of animals, contrast sensitivity of vision, and the Weber-Fechner law. Sensory Systems, 37, 46–60.
95. Binhi, V. N. (2023). Statistical amplification of the effects of weak MFs in cellular translation. Cells, 12, 724.
96. Mohler, K., & Ibba, M. (2017). Translational fidelity and mistranslation in the cellular response to stress. Nature Microbiology, 2, 17117.
97. Parker, J. (1989). Errors and alternatives in reading the universal genetic code. Microbiological Reviews, 53, 273–298.
98. Kurland, C. G. (1992). Translational accuracy and the fitness of bacteria. Annual Review of Genetics, 26, 29–50.
99. Nissley, D. A., & Jiang, Y., Trovato, F., Sitarik, I., Narayan, K. B., To, P., et al. (2022). Universal protein misfolding intermediates can bypass the proteostasis network and remain soluble and less functional. Nature Communications, 13, 3081.
100. Belyaev, I. Y., & Koch, C. B., & Terenius, O., & Roxstrom-Lindquist, & K., Malmgren, & L. O. G., Sommer, W. H., et al. (2006). Exposure of rat brain to 915 MHz GSM microwaves induces changes in gene expression but not double stranded DNA breaks or effects on chromatin conformation. Bioelectromagnetics, 27, 295–306.
101. Drummond, D. A., & Wilke, C. O. (2008). Mistranslation-induced protein misfolding as a dominant constraint on coding-sequence evolution. Cell, 134, 341–352.
102. Legrain, P., & Aebersold, R., & Archakov, A., & Bairoch, A., & Bala, K., & Beretta, L., et al. (2011). The human proteome project: Current state and future direction. Molecular & Cellular Proteomics, 10.
103. Stovbun, S. V., & Zlenko, D. V., & Bukhvostov, A. A., & Vedenkin, A. A., & Skoblin, A. A., & Kuznetsov, D. A., et al. (2023). Magnetic field and nuclear spin influence on the DNA synthesis rate. Scientific Reports, 13, 465.
104. Agliassa, C., & Maffei, M. E. (2019). Reduction of geomagnetic field (GMF) to near null magnetic field (NNMF) affects some Arabidopsis thaliana clock genes amplitude in a light independent manner. Journal of Plant Physiology, 232, 23–26.
105. Binhi, V. N. (2016). Primary physical mechanism of the biological effects of weak magnetic fields. Biophysics, 61, 170–176.
106. Dhiman, S. K., & Galland, P. (2018). Effects of weak static magnetic fields on the gene expression of seedlings of Arabidopsis thaliana. Journal of Plant Physiology, 231, 9–18.
107. Blank, M., & Soo, L., & Lin, H., Henderson, A. S., & Goodman, R. (1992). Changes in transcription in HL-60 cells following exposure to alternating currents from electric fields. Bioelectrochemistry and Bioenergetics, 28, 301–309.
108. Binhi, V. N., & Chernavsky, D. S. (2005). Stochastic resonance of magnetosomes fixed in the cytoskeleton. Biophysics, 50, 599–603.
109. Kirschvink, J. L., & Winklhofer, M., & Walker, M. M. (2010). Biophysics of magnetic orientation: Strengthening the interface between theory and experimental design. Journal of the Royal Society Interface 7, 179–171.
Результаты процедуры рецензирования статьи
Рецензия скрыта по просьбе автора
Ссылка на эту статью
Просто выделите и скопируйте ссылку на эту статью в буфер обмена. Вы можете также
попробовать найти похожие
статьи
|
|



